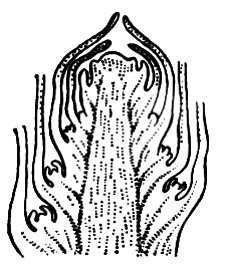
По положению различают П. верхушечные, или терминальные, за счёт которых побег нарастает в длину, и боковые, обеспечивающие ветвление и образование системы побегов. У большинства семенных растений боковые П. сидят в пазухах листьев и называются пазушными, или аксиллярными. Они закладываются как наружные меристематические бугорки (экзогенно) в пазухах листовых зачатков на ранних этапах развития материнского побега, нередко ещё внутри материнской П. (рис. 1 ). Кроющий лист, в пазухе которого сидит П., обеспечивает её питание и защиту. В пазухе листа обычно образуется одна П., хотя у некоторых растений бывают т. н. добавочные П., сидящие одна над другой (сериальные, как у жимолости) или рядом, бок о бок (коллатеральные, как в луковице чеснока). Вне пазух, эндогенно (во внутренних слоях тканей взрослого стебля, корня, а иногда — листа) закладываются т. н. придаточные, или адвентивные, П. У высших споровых растений (мхи, папоротникообразные) П. чаще придаточные.
Наружные листья П. или их части (прилистники, основание листа, часть черешка) иногда превращаются в почечные чешуи (рис. 2 ), нередко имеющие особое анатомическое строение (с сильным развитием покровных тканей, с волосками, опробковевшие или одревесневшие, выделяющие смолу и т.д.). Такие П., называют закрытыми, свойственны большинству деревьев, кустарников и многолетних трав холодного и умеренного поясов, а также субтропических и тропических областей с сухим периодом. При развёртывании П. чешуи обычно опадают, а междоузлия между ними остаются короткими и на границах периодических приростов побегов образуются группы сближенных рубцов — почечные кольца. По ним у деревьев И кустарников умеренного пояса можно определить границы годичных приростов н возраст ветвей. Открытые, или голые, П. (без чешуй) преобладают у деревьев влажнотропические леса, но часто встречаются и в умеренном поясе на незимующих, а иногда и на зимующих побегах трав (например, у живучки и элодеи). У древесных растений умеренных широт открытые П. можно видеть весной в период «открытого роста» на концах молодых побегов; зимующие открытые П. встречаются у них очень редко (например, у крушины ломкой).
Расположение листьев в П, по отношению друг к другу называется почкосложением, или почкосмыканием (рис. 3 ). В зависимости от того, как сложена (и сложена ли вообще) пластинка каждого листа, в П. различают типы листосложения (рис. 4 ). Почкосложение и листосложение — наследственные признаки, учитываемые в систематике растений.
Как правило, вновь образующаяся П. развёртывается в побег не сразу, а только после заложения в ней определённого числа листовых зачатков (зрелая П.). Часто зрелая П. переживает более или менее длительный период покоя, связанный с неблагоприятным временем года (зима или сухой период); такие П., дающие побеги весной или с наступлением влажного сезона, называют П. возобновления. В зимующих П. возобновления осенью может быть сформирован весь побег будущего года, включая соцветия (сирень, бузина, ландыш, копытень) или только часть его (липа, золотая розга); у многих деревьев и кустарников зимующие П. четко делятся на вегетативные и цветочные (берёза, яблоня, тополь).
П. могут закладываться в пазухах всех листьев, и от этого зависит способность растений к ветвлению и возобновлению; в результате у деревьев и кустарников значительная часть П. не развёртывается сразу; они называются спящими П. и сохраняют жизнеспособность в течение многих лет. Такие П. развёртываются при повреждении вышележащих частей ствола и ветвей; из них образуются водяные побеги , часто в виде пнёвой поросли. У кустарников спящие П. регулярно дают новые стволики, сменяя прежние. Способность спящих П. давать побеги широко используется при формировании кроны у декоративных и плодовых деревьев, при разведении живых изгородей и т.д. Из придаточных П. на корнях образуются надземные побеги — корневые отпрыски. Роль П. в вегетативном размножении растений очень велика (рис. 5 ). У некоторых растений особые, т. н. выводковые П. развиваются в пазухах листьев, в соцветиях, даже на листьях (бриофиллюм); они отпадают от материнского растения, прорастают, укореняются и дают новую особь (см. Живородящие растения ).
В плодоводстве при вегетативном размножении путём прививки (окулировки) используют небольшой участок стебля с покоящейся П., называют глазком. Это название применяется и для П. на клубнях картофеля.
Лит.: Серебряков И. Г., Морфология вегетативных органов высших растений, М., 1952; Синнот Э., Морфогенез растений, пер. с англ., М., 1963; Леопольд А., Рост и развитие растений, пер. с англ., М., 1968.
Т. И. Серебрякова.
Рис. 3. Различные типы почкосложения: а — створчатое (листья соприкасаются краями); б — черепитчатое (листья прикрывают друг друга краями); в — прикрывающее, или скрученное (у каждого листа один край прикрыт краем соседнего листа); г — полуприкрывающее (из 5 листьев мутовки 2 не прикрыты, 2 прикрыты с обеих сторон, 1 с одной стороны прикрыт, а с другой — сам прикрывает соседний); д — объемлющее (каждый лист охватывает все листья, лежащие внутрь от него).
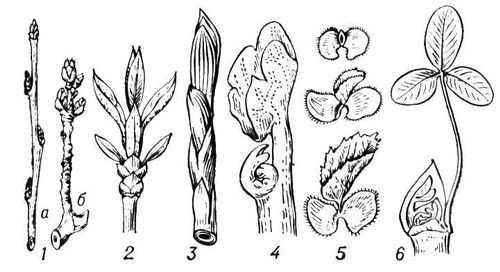
Рис. 5. Почки, служащие для вегетативного размножения: 1 — верхушечная почка на столоне картофеля; 2 — то же на усе земляники; 3 — придаточные почки на корне вязеля; 4 — зимующая подводная почка стрелолиста.
Рис. 4. Различные типы листосложения в почках: а — складчатое; б — многократно-складчатое; в — свёрнутое; г — отвёрнутое; д — завёрнутое.
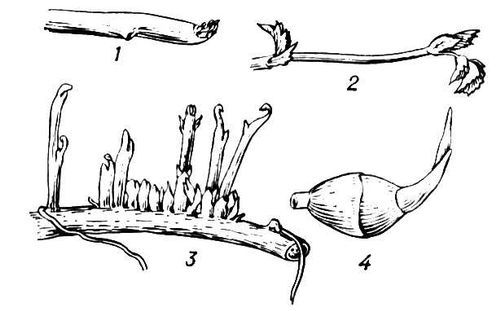
Рис. 2. Типы почек: 1 — закрытые почки осины (а — вегетативные, б — цветочные); 2 — начало развёртывания почки сирени (видны почечные чешуи и переходные листья); 3 — начало развёртывания почки чемерицы (видны почечные чешуи); 4 — почечные чешуи лещины (образованы прилистниками); 5 — открытая почка на верхушке побега настурции; 6 — схема верхушечной почки побега клевера (роль чешуй выполняют прилистники).
Рис. 1. Почка побега семенного растения (продольный разрез — схема); в пазухах зачатков нижних листьев видны зачатки пазушных почек.
Почки
По'чки, важнейшие парные органы выделения позвоночных животных и человека, участвующие в водно-солевом гомеостазе , т. е. в поддержании постоянства концентрации осмотически активных веществ в жидкостях внутренней среды (см. Осморегуляция ), постоянства объёма этих жидкостей, их ионного состава и кислотно-щелочного равновесия . Через П. выводятся из организма конечные продукты азотистого обмена, чужеродные и токсические соединения, избыток органических и неорганических веществ. П, участвуют в метаболизме углеводов и белков, в образовании биологически активных веществ, регулирующих уровень артериального давления, скорость секреции альдостерона надпочечниками и скорость образования эритроцитов.
Сравнительная морфология . В процессе эволюции позвоночных животных происходила последовательная смена трёх типов П.: предпочки, или головной П. (пронефрос ), первичной, или туловищной, П. (мезонефрос ), вторичной, или тазовой, П. (метанефрос ). Эти типы П. сменяют друг друга и в течение индивидуального развития высших позвоночных. Все типы П. развиваются из ножки сомита — нефротома . Первичная П. — орган выделения, функционирующий в послезародышевый период у круглоротых, рыб, земноводных, молодых ящериц и у некоторых млекопитающих (клоачные, сумчатые) до наступления у них половой зрелости. У всех остальных позвоночных и у человека первичная П. сменяется вторичной ещё у зародышей. Основная морфо-функциональная единица П. — нефрон . В процессе эволюции П. у позвоночных происходило развитие структур нефрона и их видоизменение, связанное с приспособлением животных к различным средам. У круглоротых (миноги) все нефроны начинаются из общей полости гломуса, в которую происходит ультрафильтрация из кровеносных капилляров. У рыб и остальных позвоночных имеется уже мальпигиево тельце , которым начинается каждый нефрон. Только у некоторых видов морских костистых рыб нефроны не имеют клубочков. В П. всех позвоночных нефрон имеет проксимальный сегмент, а у подавляющего большинства животных (кроме нескольких видов морских костистых рыб) и дистальный. У птиц и млекопитающих развивается новая морфология, структура — петля Генле, канальцы которой служат основным элементом мозгового вещества П.