Папоротник женский
Па'поротник же'нский , кочедыжник женский (Athyrium filix-femina), травянистое многолетнее растение семейства кочедыжниковых. Пластинка листа дважды-триждыперистораздельная. Сорусы эллиптические серповидно-изогнутые или крючковидные, расположенные вдоль боковых жилок сегментов листовой пластинки. Индузий с надрезным краем, по созревании сбрасывающийся. П. ж. распространён в лесной зоне Северного полушария, растет в сырых тенистых местах. Лекарственное (глистогонное) и декоративное растение.
Папоротник женский; внизу — сегмент с сорусами.
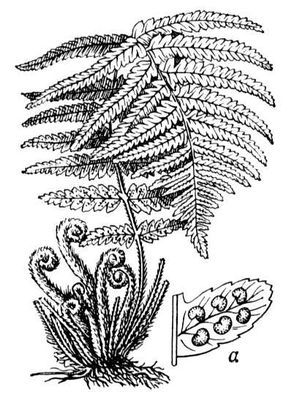
Папоротник мужской
Па'поротник мужско'й , щитовник мужской (Dryopteris filix-mas), травянистое многолетнее растение семейства аспидиевых. Пластинка листа дваждыперисторассечённая, черешок и корневище с крупными тёмными чешуями. Сорусы с почковидным индузием расположены на боковых жилках сегментов листовой пластинки. Растет в лесной зоне Евразии и Северной Америки, в тенистых местах по кустарникам. Экстракт корневища применяют как глистогонное против ленточных глистов и остриц: в корневищах П. м. содержатся вещества (производные филициновой кислоты), вызывающие паралич мускулатуры у паразитов кишечника.
Папоротник мужской; а — сегмент листа с сорусами.
Папоротники
Па'поротники (Polypodiophyta), отдел высших растений, занимающий промежуточное положение между риниофитами и голосеменными. От риниофитов П. отличаются главным образом наличием корней и листьев, а от голосеменных — отсутствием семян. П. произошли от риниофитов, к которым древнейшие девонские П. были очень близки (некоторые наиболее примитивные роды были по существу промежуточными формами между риниофитами и типичными П.). Для П., как и для других высших растений, характерно чередование поколений — бесполого (спорофита ) и полового (гаметофита ), с доминированием бесполого поколения. Спорофит П. — травянистое или древовидное растение большей частью с крупными, многократно рассеченными листьями (молодые — обычно улиткообразно свёрнутые). Листья П. произошли из дихотомических ветвлений риниофитов в результате их уплощения и ограничения в росте с последующей дифференциацией верхней и нижней поверхности листовой пластинки. Это подтверждается длительным верхушечным ростом, крупными размерами и сложнорассечённой формой листьев девонских П. Стебли древнейших девонских П. мало отличались от типичных стеблей риниофитов и имели проводящую систему типа протостелы, которая сохранилась у некоторых современных П. (например, у схизеевых и многих глейхениевых). У большинства современных П. проводящая система представляет собой сифоностелу или чаще диктиостелу (см. Стела ). Большинство П. лишено способности ко вторичному росту стебля. Ксилема обычно состоит из трахеид, но у орляка и марсилеи есть также настоящие сосуды. У П. имеются придаточные корни, которые, вероятно, произошли из видоизменённых подземных корневищеподобных органов (ризомоидов) риниофитов. Для П. характерно большое разнообразие формы, внутреннего строения и размеров. Листья их варьируют от многократноперисто- рассечённых до цельных, от гигантских — длиной 5—6 м (у некоторых представителей мараттиевых и циатейных) и даже до 30 м (вьющиеся листья у Lygodium articulatum из семейства схизеевых) до крошечных листьев длиной всего 3—4 мм , состоящих из 1 слоя клеток (у Trichomanes goebelianum из семейства гименофилловых). Длина стеблей у П. варьирует от нескольких сантиметров до 20—25 м (у некоторых видов рода циатея). Они бывают подземными (корневища) и надземными, прямостоячими и вьющимися, простыми и ветвистыми. У большинства спорангии расположены на обычных зелёных листьях; у некоторых листья дифференцированы на спороносные (спорофиллы) и вегетативные, зелёные (или же дифференцированы сегменты одного и того же листа, как, например, у чистоуста). Первоначально спорангии сидели на верхушках сегментов листа — то есть терминально; по мере уплощения дихотомически разветвленного листа они переместились на его края, заняв маргинальное положение. Это характерно, например, для семейства гименофилловых и диксониевых. У большинства современных П. спорангии расположены на нижней (абаксиальной) стороне листа. Первоначально спорангии сидели на листьях поодиночке (как у современного семейства схизеевых), но в процессе эволюции они группировались в сорусы — голые у примитивных П. (мараттиевых, чистоустовых, глейхениевых) или имеющие защитный покров — индузий (у большинства современных П.). У некоторых родов семейства мараттиевых спорангии в сорусе срослись между собой, образовав так называемые синангии. Для примитивных П. (ужовниковые, мараттиевые) характерны спорангии относительно крупные, с многослойной стенкой и большим количеством спор; у большинства современных П. они мельче, с однослойной стенкой и производят небольшое число спор. Большая часть ныне живущих П. имеет механизм для раскрывания спорангия (обычно в форме кольца).
Большинство П. — равноспоровые; лишь в некоторых линиях их эволюции уже в палеозое возникли разноспоровые, у которых споры дифференцировались на микроспоры и мегаспоры . Среди современных П. к разноспоровым относятся лишь 3 небольших семейства водных П.: марсилеевые, сальвиниевые и азолловые. У равноспоровых П. спора при прорастании даёт начало обоеполому гаметофиту, который у большинства П. представляет собой нежную зелёную недолговечную пластинку более или менее сердцевидной формы, с рассеянными на её поверхности (преимущественно нижней) половыми органами — архегониями и антеридиями. У разноспоровых П. гаметофиты однополые и сильно редуцированы. Микроспоры дают начало крошечному мужскому гаметофиту, мегаспоры — несколько более крупному женскому гаметофиту. Сперматозоиды у П. многожгутиковые, как у саговников и гинкго . Вегетативно П. размножаются корневищами, а также при помощи появляющихся на листьях выводковых почек.
Отдел П. включает 1 класс Polypodiopsida, подразделяемый на 8 подклассов, в том числе 3 вымерших в девоне — перми: Protopteridiidae (ближе всего стоявший к риниофитам), Archaeopteridiidae и Noeggerathiidae. Современные П. представлены 300 родами (около 10 000 видов). Из подклассов, представленных в современной флоре, наиболее примитивны семейство Ophioglossidae (ужовниковые), включающее 70 видов из 4 родов, и Marattiidae (около 190 видов из 6 родов). Самый обширный подкласс современных П. — Polypodiidae (Filicidae), включающий около 300 родов (до 10 000 видов). Большинство семейств этого подкласса неизвестно ранее позднего триаса, но чистоустовые встречаются уже в перми, а схизеевые даже с карбона. С позднего триаса бурно развиваются уже почти все семейства этого подкласса: многие из них достигли расцвета в раннемеловую эпоху. Представители Polypodiidae произрастают во всех частях света, но главным образом во влажных тропиках и субтропиках, составляя важный элемент растительности, особенно влажных ущелий и горных лесов. В умеренном поясе они растут чаще всего в тенистых лесах, глубоких ущельях и на болотах. Некоторые из них — ксерофиты , растут на скалах или на склонах между камнями. Другие виды растут в воде, немногие — в составе мангровой растительности. В тропических странах очень много эпифитных П., главным образом из семейства гименофилловых, многоножковых и некоторых др. Представители семейства циатейных — преимущественно древовидные П., часто с очень высокими стволами (до 20—25 м ), увенчанными кроной больших дважды- или триждыперистых листьев. Самый большой род циатея включает свыше 600 видов, произрастающих главным образом в горах во влажных тропиках, а также в Чили, Новой Зеландии и Южной Африке. Некоторые новозеландские виды растут у ледника Франца-Иосифа на Южном острове и проникают даже на субантарктические Оклендские острова; диксония антарктическая достигает в Тасмании 45° южной широты. Во флоре СССР этот подкласс представлен семейством чистоустовых, гимепофилловых, птеридовых, асплениевых, аспидиевых, дербянковых, многоножковых и плагиогириевых. От равноспоровых Polypodiidae произошли 2 подкласса — Marsileidae (1 семейство марсилеевых, представленное в СССР родами марсилея и пилюлария) и Salviniidae (2 семейства — сальвиниевые и азолловые; первое представлено в СССР единственным в семействе родом сальвиния). Оба эти подкласса — водные растения: марсилеевые прикрепляются корнями к субстрату, спороносная часть листа превращена у них в замкнутое вместилище сорусов (спорокарпий); сальвиниевые и азолловые плавают на поверхности воды и сорусы у них сидят свободно на погруженных в воду листьях (сальвиния) или на погруженном в воду сегменте двураздельного листа (азолла).