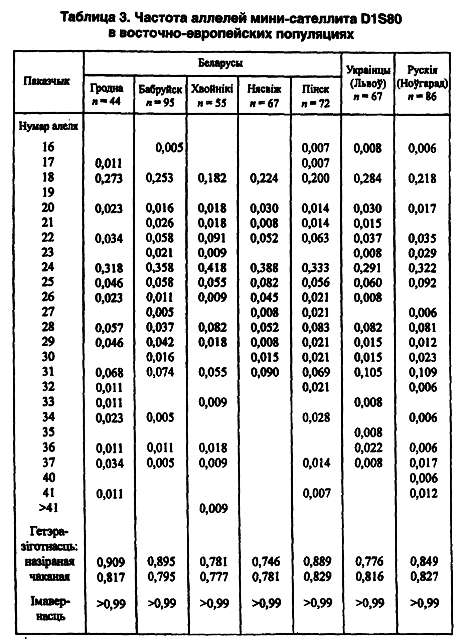
Однако на карте распространения 5-го аллеля локуса DM (карта 10) в восточнославянских популяциях выделяются 4 геногеографических региона с вектором северо-запад — юго-восток (Лимборская С. А. и др., 2002). Согласно данным указанной публикации, наблюдаемая и ожидаемая величины гетерозиготности беларуских популяций весьма близки друг другу, без отклонения от равновесия Харди-Вайнберга. И последнее замечание по данному локусу. Самые редкие аллели (17-й и 33-й) выявлены только у сельских (до 4-го колена и далее) беларусов и коренных немцев — у 0,7 и 0,5 процента соответственно. Все это косвенно свидетельствует о возможном воздействии стабилизационного отбора.
С целью решения задач этнической антропологии мы исследовали полиморфизм еще двух маркеров ДНК — SCA1 и CCR5. Из восточноевропейских этносов наряду с беларусами были обследованы башкиры и удмурты в первом случае, русские и башкиры — во втором. Изучались гетерозиготность и концентрации генотипов и аллелей. Наш этнос отличается от двух уральских максимальными частотами (10 % и более) гетерозиготных генотипов 22/27, 19/22, 19/28, а также 22/24, и гомозиготы 22/22 (в пределах 7–5 %). Гомозигота 19/19 у нас вообще не выявлена, тогда как среди представителей монголоидной расы она достигает 4 процентов. Для 13 аллелей локуса СА1 теоретически должен существовать 91 вариант генотипов. Но если среди волго-уральских популяций выявлена почти половина их, то в Беларуси — только четверть.
Популяции всех трех изучавшихся этносов по наиболее распространенным аллелям (19-й, 22-й, 27-й) статистически значительно различаются между собой. Показатель общей гетерозиготности наиболее возрастает от европеоидов (0,72) в сторону негроидов (0,86) (Лимборская С. А. и др., 2002).
Согласно результатам исследования по анализу генофонда маркера CCR5 среди беларуских, русских и башкирских популяций, его информационная значимость тоже достаточно высокая. Во всех изученных выборках распределение генотипов полностью соответствует закону Харди-Вайнберга (р<0,05). Концентрации всех генотипов и аллелей у беларусов и башкиров полярные, а у русских — промежуточные между ними. Здесь межэтнический вектор изменчивости имеет, согласно С. Лимборской, широтное направление. Аллельные варианты этого маркера по своим концентрациям существенно различаются в популяциях разных рас. Происходит это преимущественно за счет новых мутаций среди северных европейцев с последующим распространением на юг. Однако не исключена также связь с иммунитетом или с адаптационными возможностями как отдельных организмов, так и популяций в целом (Лимборская С. А. и др., 2002).
Более основательно исследована популяционно-генетическая особенность микросателлитного локуса DM гена миотанинпротеинкиназы (хромосома 19), являющегося причиной такого неврологического заболевания, как миотоническая дистрофия. На первый взгляд, нет существенных различий в распределении аллельных вариантов у разных территориальных групп беларусов по анализировавшимся выборкам (Попова С. Н., Микулич А. И. и др., 1999). Но только на первый взгляд. Локус DM в обычных условиях ведет себя нейтрально и поэтому становится пригодным для изучения популяций. Количество аллельных вариантов в разных популяциях беларусов неодинаково: от 9 до 12 из возможных 17.Что существенно — у беларусов выделены 4 аллельных повтора, пока еще не открытых в Европе: 19-й, 25-й, 27-й, 29-й. Однако такое явление не привело к отклонению равновесия Харди-Вайнберга (р< 0,89–0,99) и нет существенного различия (не более 2 %) между теоретической и наблюдаемой гетерозиготностью.
Выявленное бимодальное распределение повторов с пиками 5-го и 11–13-го аллелей фактически одинаковое как в беларуской популяции, так и в общеевропейской. Однако наличие у беларусов четырех перечисленных редких аллелей дополнительно свидетельствует об их древней автохтонности. К такому выводу мы пришли на основе подсчетов генетических расстояний между выборками с помощью пакета программ PopGene на базе алгоритма, предложенного М. Неем в 1978 году.
На соответствующей дендрограмме гродненская и хойникская популяции занимают полярные позиции, а Несвижская и бобруйская формируют промежуточный кластер. Почти меридиональный вектор изменчивости снова сохраняет свое географическое направление. Картографическая модель распределения в географическом пространстве 5-го аллеля как супермажорного (41 + 3) полностью подтверждает наш предыдущий аналитический прогноз 2002 года. Популяционные характеристики соседних этносов плавно перетекают от минимума у финскоязычных этносов к максимуму у московских русских и молдаван.
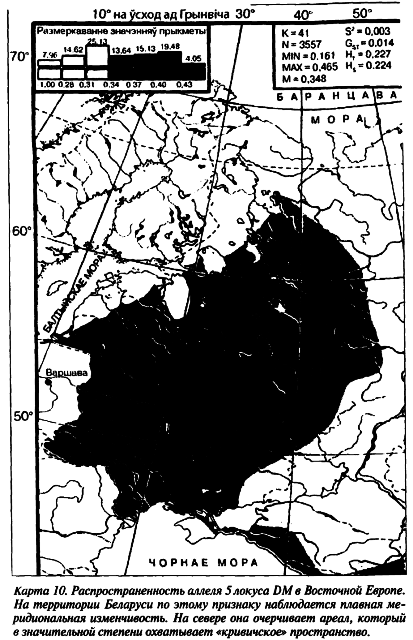
На геногеографической карте аллеля 11 локуса DM его минор[109] находится в центральной части Беларуского Полесья, а смежные концентрации объединяют южных беларусов с жителями Украинского Полесья и восточных беларусов с населением смежных регионов Российской Федерации (карта 11). Аллель 13 обладает противоположной частотной изменчивостью.
Согласно данным палеоантропологии, предки с этих территорий имели наибольшее число европеоидных признаков (Алексеев В. П., 1969). Археологи, в свою очередь, свидетельствуют о распространении здесь древностей зарубинецкой культуры (Третьяков П. Н., 1966; Седов В. В., 1982). Остается, правда, открытым вопрос: чьими предками могли быть носители зарубинецкой культуры — славян или балтов? Наш антропологический и популяционно-генетический материал свидетельствует в пользу тех и других.
Распределение аллеля 14 и гетерозиготности по всему локусу DM на беларуской территории полностью соответствует ранее выделенному нами по антропогенетическим особенностям географическому направлению с юго-запада на северо-восток (карта 12). Наш суммарный анализ частот редких аллелей (от 20 и выше) показал, что их концентрация максимальна именно в беларуской популяции. Таким образом, этот пик, который до недавнего времени локализовали в Центральной Европе, переместился на территорию Беларуси. Этот факт еще раз демонстрирует необходимость и важность изучения антропологических и генетических признаков не только по их средним показателям, но и обязательно (как предлагает Ю. Г. Рычков) с учетом даже чрезвычайно редких признаков. То обстоятельство, что на частотный спектр аллелей локуса DM действительно влияют не только монголоидно-европеоидные соподчинения, но и естественный отбор (с последующим закреплением отдельных аллелей на конкретных территориях), достаточно аргументировано (Лимборская С. А. и др., 2002).
Наш предыдущий анализ полиморфизма триплетного повтора гена миотанинпротеинкиназы DM выявил 17 аллельных его вариантов с числом единиц от 5 до 29. Аллели с 6-м и 7-м повторами вообще не выявлены. Уровень гетерозиготности колеблется в разных популяциях от 71 до 81 процента, что статистически достоверно не отличает его от ожидаемого — 73–79 процентов (индекс фиксации — 0,002) и косвенно указывает на фактическую селекционную нейтральность нормального полиморфизма STG-повторов гена DM среди беларуских популяций.
Как видим, концентрации каждого из аллельных вариантов ДМ имеют свою особенность распространения среди коренных беларусов. Поэтому снова возникает потребность обращения к обобщенному массиву информации с помощью метода главных компонент частотных показателей посредством многомерного статистического анализа. Он помогает выявить общие закономерности изменчивости. Например, территория современной Беларуси находится в промежуточной зоне, где первая главная компонента определяет регион равновесия двух противоположных тенденций (Лимборская С. А. и др., 2002). В частности, северо-восточная и юго-западная провинции Беларуси имеют очевидную связь с соответствующими прилегающими территориями соседних стран.