С другой стороны, имеются ли основания, как это делают московские авторы, соотносить восточнославянский ареал с территорией расселения венедов и антов? Трудно сказать. Возможно, и есть. Но требуются дополнительные доказательства.
В монографии «Этногеномика и геногеография народов Восточнгой Европы» (2002 год) отмечено, что достаточно полно изученный генофонд Беларуси использован в качестве модельного объекта наряду с Черноморско-Балтийским и Северо-Евразийским регионами (Лимборская С. А. и др., с. 25). Цитируем:
«Каждый последующий генофонд как минимум на ранг крупнее предыдущего и позволяет проследить закономерности более высокого порядка. С другой стороны, выявленные закономерности полностью распространяются и на генофонды более низких уровней.
Такой принцип позволяет выявить общее и особенное в пространственной структуре каждого из генофондов, а с методической точки зрения дает возможность провести анализ с внутренним контролем».
Унифицированные методики сбора и обработки экспедиционных материалов помогли выявить главные компоненты генотипной изменчивости путем усовершенствования известной методологии их картографирования. Так была создана серия унифицированных карт генетических ландшафтов, конденсирующих в себе основные параметры изменчивости частот генов и основную часть общей дисперсии. Этим способом уже проверена достоверность выявленных закономерностей изменчивости генофонда сразу по четырем независимым системам данных — из антропологии, дерматоглифики, классической и молекулярной антропогенетики (Балановский О. П. и др., 2000, 2001).
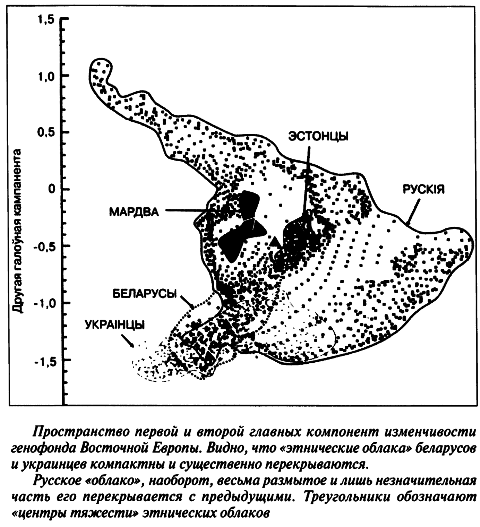
Все названные обстоятельства, по нашему мнению, позволяют — с учетом знаний об истории изученных этносов — собственным взглядом увидеть, как наши беларуские популяции распределены в геногеографическом пространстве главных компонент. Так, центры тяжести «этнических облаков» беларусов и украинцев (по терминологии Е. П. Балановской) в пространстве двух главных компонент размещаются рядом, но сами «облака» перекрываются только наполовину, едва касаясь «облака» русских (см. рисунок на с. 152).
При этом, если украинский этнос вообще не граничит с финно-угорской компонентой, а беларуский — лишь соприкасается с ней, то «русские популяции во всех диаграммах оказываются в одном кластере с финно-угорскими, а не славянскими этносами» (Лимборская С. А. и др., 2002).
Следовательно, это еще один аргумент в пользу исторически и экологически зависимых особенностей каждого из восточнославянских этносов. В последнее время археологи все чаще сравнивают результаты своих исследований с нашими и находят соответствующие параллелли (Штыхов Г. В., 2002). Следует считать не только желательным, но и необходимым сравнение археологических материалов с материалами геногеографии. Такое сравнение выявляет специфику генофонда, которая посредством механизма антропогенетической преемственности дошла от древности до современности.
Кстати можно упомянуть недавно изданную на родине книгу Витовта Тумаша «Избранные труды», в которой напечатана его статья середины XX века о балтском элементе в этногенезе беларусов. Сылаясь на своих предшественников-антропологов, считавших беларусов «наичистейшим славянским народом, наилучшим образом сохранившим свои славянские особенности», автор объясняет: «такой взгляд сложился прежде всего потому, что в противоположность русским и украинцам наши пращуры слились не с чужими, антропологически далекими от них финскими или монгольскими элементами, а с расово близкими индоевропейскими балтами».[105]
Обратимся к результатам картографирования не главных компонент, а генетических расстояний.[106] Авторы указанной публикации, учитывающие и наши данные по классическим генным маркерам, строят карты с принципиально новым генетическим рельефом, где отмечают расстояние от любой среднерегиональной (реперной) частоты. Однако на карте генетических расстояний от средних русских частот генов (см. карту 9), по их собственным словам, «основная площадь Восточной Европы окрашена в светлые тона, указывающие на общность с русским генофондом». Тем самым фактически игнорируется присутствие своеобразного беларуского генофонда.
Согласиться с таким тезисом нельзя. Генетический и морфологический материал свидетельствуют об отличии беларуского этноса в географическом пространстве и историческом времени. Напомним, что еще в середине 50-х годов XX века В. В. Бунак, изучая в рамках расоведения физический тип беларусов, предлагал этногенетическую гипотезу о наличии в их фенотипе очень древней (с эпохи мезолита) северо- и южноевропеоидной примеси (Бунак В. В., 1956). Эта идея нашла подтверждение в позднейших исследованиях (Битов М. В. идр., 1959; Дьяченко В. Д., 1965; Денисова Р. Я., 1975).
На карте генетических расстояний населения Восточной Европы от средних беларуских частот генов (см. карту 7), построенной на основе значительного количества маркеров (57 аллелей 21 локуса), прекрасно видна своеобразная особенность генофонда беларусов. К ним присоединяются коренные жители Псковской, Новгородской, Смоленской и Брянской областей, Виленского края, Украинского Полесья.
По нашему мнению, тем самым зафиксированы исторические явления не XVII–XVIII веков, а на три тысячи лет раньше. С учетом всего изученного материала ареал беларуского генофонда в общих своих очертаниях соответствует границам Великого Княжества Литовского середины XV века. Многовековое существование этой беларуской державы, единство происхождения, своеобразие языка, духовной и материальной культуры беларусов способствовали консолидации древнего этноса.
ГЛАВА 8.
Этногеномика популяций
Восточной Европы по ДНК-маркерам
До недавнего времени, изучая этническую геногеографию, мы пользовались преимущественно многочисленными выборками индивидуальных геномов. Диктовалось это сравнительно малым числом исследованных локусов. Нынешний молекулярно-биологический подход к изучению генетико-популяционной дифференциации позволяет значительно расширить количество геномных групп. Он существенно увеличивает достоверность полученных результатов по причине более эффективного нивелирования влияния возможных факторов микроэволюции. Последние имеют различную значимость как для отдельных генов, так и для отдельных популяций. Селекция человека как социального организма в историческое время генетически могла оставаться (и, скорее всего, оставалась) генетически нейтральной.
Все эти обстоятельства фактически свидетельствуют об одинаковой роли обобщенных генов при дифференциации генофондов по совокупности классических факторов.
«Исследование генофонда, то есть совокупности всех генов в популяции, является собственно популяционной задачей, при разрешении которой проблема функции гена второстепенна. Поэтому на завершающем этапе исследования не только возможно, но и необходимо получить обобщенные популяционно-генетические характеристики генов в системе генофонда».[107]
Именно с такими характеристиками мы и будем иметь дело в разделе этногеномики. Имеются в виду уровни генетической межпопуляционной дифференциации (Dst) и гетерозиготности популяций (Hs). Гетерозиготное различие между регионами как раз и отражает историко-культурные условия прошлого и демографические особенности населения соответствующих территорий.
В своих предыдущих публикациях мы широко пользовались также статистической категорией обобщенного гена — так называемым генетическим расстоянием между генофондами — d (квадратичное), которое, между прочим, весьма близко к среднему Fst (Edwards A. W., Cavalli-Sforza L. L., 1972). Связь действительно вполне очевидная. Она точно фиксирует усиление генетического разнообразия между популяциями соответствующим увеличением генетического расстояния между ними. Эти параметры достаточно объективны и линейно пропорциональны времени их дивергенции от популяций предков.