Однако точность этого соответствия будет наиболее высокой при том условии, что эффективно-репродукционные размеры популяций (N) остаются если не стабильными, то хотя бы без значительного (свыше четверти) притока иммигрантов (М). (Шереметьева В. А., 1975).
Уже предварительный анализ полиморфизма аллелей и гаплотипов позволил получить статистические результаты, отследить геногеографию обобщенных характеристик. Все это, вместе взятое, способствует дальнейшему поиску путей сохранения генодемографического разнообразия и возможной адаптивности.
В рамках отдельного беларуско-российского проекта ФФД № Г99Р-02 выполнен предварительный анализ по локусам DM, SCA1, IGHI, D1S80. Всего обследовано 8 выборок. Во всех популяциях суммарно выявлены практически все генотипы, характерные для европеоидной расы генотипа локуса 1GHI. Однако наиболее редкие из них встречаются в Иванове (10/11, 10/14, 11/12), Пинске (10/17, 7/10, 16/17), в Молодечно помимо двух последних еще и 8/17, в Мяделе 12/15, Климовичах 11/12. Из всех аллелей 14-й имеется только в Иванове, 7-й в Молодечно и Пинске, 15-й в Полоцке и Городке. 13-й аллель отсутствует во всей Центральной и Восточной Европе. Кроме того, у этнических беларусов отсутствует 11-й аллель. Пять аллелей (8-й, 9-й, 10-й, 12-й, 16-й) выявлены во всех изученных выборках.
На примере предыдущих материалов по изосерологической и морфофункциональной изменчивости нами установлен факт достаточно устойчивой генетической памяти в локальных сельских популяциях. По количеству 10-го и 12-го иммунноглобулиновых аллелей существует так называемое бимодальное распределение с модальной их частотой 0,71 + 0,03, в своеобразном виде — в границах экологических макровыборок: от 0,678 ± 0,03 на юге до 0,748 ± 0,01 на севере. Так, наибольшее число аллелей отсутствует в Придвинье — 7-й, 11-й, 14-й и 17-й; в Центральной Беларуси — 11-й, 14-й и 15-й, на Полесье — только два (11-й и 15-й).
Максимальная палитра изменчивости выявлена опять в Полесской геоморфологической провинции. Это явление — реликтовые особенности данной провинции — соотносится с нашими предыдущими открытиями редких гаплотипов в резус-системе. Матрица и дендрограмма локальных популяций сохраняют ту же меридиональную тенденцию. Видимо, в этом случае можно считать, что наряду с устойчивой генетической памятью существует и экологическая зависимость прапопуляций в процесе эволюции. Не исключено также миграционное влияние.
Таким образом, положено начало формированию соответствующей базы данных других аллелей, в частности, ответственных за иммуноглобулиновые факторы. Пока по этому локусу обследованы популяции трех сельских районов Минской области и трех средних городов (Молодечно, Полоцка, Пинска) в трех экологических провинциях Беларуси: Центральной, Придвинской и Полесской. Каждая из изученных популяций имеет свой характер распределения частот встречаемости аллелей IGHJ (HVR-Ig) (см. таблицу 1).

В настоящее время по данным литературы известно до восьми аллелей в разных этническо-расовых популяциях мира. Среди беларуских популяций идентифицированы все 8, но каждый из них имеет свои особенности. Так, в молодечненской и пинской популяциях не выявлен 15-й аллель, в полоцкой — 7-й и 17-й. Даже на этапе предварительного статистического анализа выявляется своя география распределения аллелей, по которой северные беларусы отличаются от южных и центральных. Наиболее часто встречаются аллели 10 и 12, что полностью совпадает с материалами литературных источников о русском и западноевропейском этносах. Это, пожалуй, то общее, что роднит европеоидов.
Особенности всегда выделяются по редким аллелям и генотипам. Например, генотипы 9–10, 9–12, 9–16 и 16–16 (4 из 19) не обнаружены в молодечненской популяции, а 8–9, 8–17 и 12–17 (3 из 19) отсутствуют в пинской популяции. Последняя в сравнении с остальными дополнительно выделяется почти вдвое более высокой концентрацией аллеля 16 (0,174). По наиболее редким гомозиготам 8–8 и 16–16 пинчуки тоже демонстрируют свое отличие. Если вспомнить выявленные нами раньше реликтовые гаплотипы резус-отрицательности — эпохи палеоевропеоидной расы в Беларуском Полесье — то подтверждается тезис об автохтонности полешуков с времен позднего неолита. Тем более, что степень гетерозиготности среди них значительно меньше даже по сравнению с населением центральной провинции (0,69 и 0,76).
Как видим, анализ даже одного высокополиморфного микросателлитного локуса предоставляет интересные данные о взаимозависимости между разными территориальными группами беларусов и позволяет проследить возможные миграционные потоки. Для получения более полной информации необходимо увеличение количества анализируемых локусов до 3–4. При высокой информативности они заменяют большинство классических маркерных систем (таких, как эритроцитные группы крови и белки кровяной плазмы). Только в таком случае ускоряется выполнение работ по анализу геногеографии в соответствии с экологическим районированием и современной экологически опасной ситуацией. С другой стороны, генотипоскопия на уровне индивида позволяет включить последнего в группу риска того или иного заболевания.
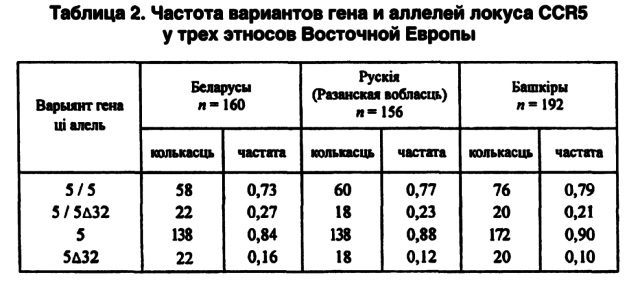
Одновременный анализ популяционных данных по обобщенным показателям поможет определить степени родства моноэтнических группировок (в границах Республики Беларусь), а также с популяциями смежных территорий. В этом смысле, как нам кажется, стоит еще раз обратить внимание на выполненное нами совместно с московскими коллегами сравнение по аллелям и генотипам локусов СА1 и CCR5 беларусов с рязанскими русскими и башкирами (Сломинский П. А. и др., 1997; Микулич А. И. и др., 1999). Из 14 аллелей локуса СА1 одинаково по 3 отсутствуют в популяциях и Беларуси, и Башкирии, причем два из них (30-й и 31-й аллели) не выявлены у обоих этносов. Кроме того, у беларусов отсутствует 32-й аллель, а у башкиров — 25-й. По частотам аллелей и генотипов гена CCR5 оказывается, что беларусы и башкиры наиболее отдалены друг от друга. Русские занимают в этом геногеографическом пространстве промежуточное положение (см. табл. 2).
Таким образом, изучение каждой новой молекулярно-генетической системы при геногеографическом подходе к анализу популяционных данных (микроэволюционных процессов и истории формирования регионального населения) расширяет возможности современной антропогенетики. Именно геногеографическая интерпретация позволила В. В. Бунаку построить интерполяционные карты АВ0-распределения Центрально-Восточной Европы (Бунак В. В., 1969). Ю. Г. Рычков создал антрополого-генетическую школу (В. А. Шереметьева, В. В. Жукова, Е. В. Балановская, А. И. Микулич и другие), усилиями которой было начато издание многотомного фундаментального труда «Генофонд и геногеография народонаселения».[108]
Приведенные в первых двух томах гипотезы и суждения нашли свое подтверждение еще по одной из исследованных нами микросателлитных систем ДНК — локусе D1S80. Мы снова обращаем внимание на геногеографические особенности распределения полученных характеристик. Например, по наличию самых редких генотипов и аллелей значительно выделяется западнополесская популяция, где их выявлено соответственно 16 и 4, тогда как в центральнобеларуской — 8 и 2, в придвинской — 4 и 0. Как видим, и эта система не противоречит нашей концепции антропогенетической изменчивости беларуского населения в направлении с юга на север или наоборот.
Подсчеты для всей макропопуляции беларуского этноса показали наличие бимодальности распределения аллелей преимущественно за счет 24-го и 18-го (0,39 ± 0,03 и 0,26 + 0,03), что дает 2/3 всего массива данных. Из 21 выявленного аллеля еще только 4 имеют 5–7-процентную величину (31-й, 22-й, 28-й и 25-й). Встречаемость каждого из остальных 15 менее одного процента (табл. 3).