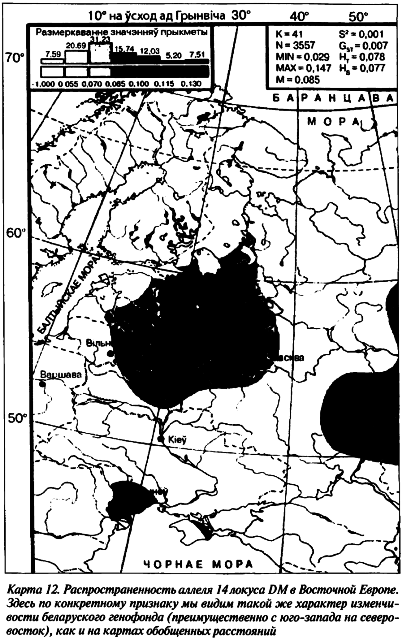
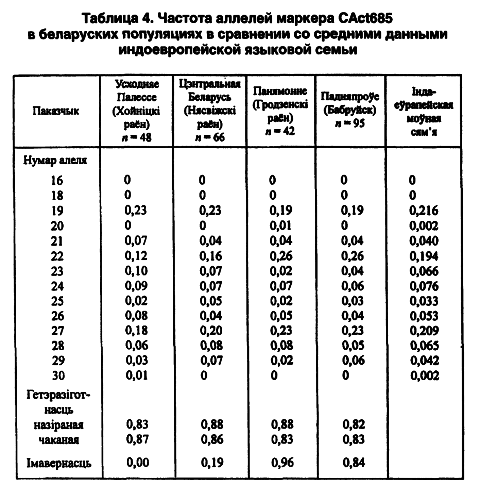
Беларуские популяции изучены еще по одному микросателлитному локусу Cact685. Генетический анализ пока завершен в 4 выборках общей численностью 241 персона. Всего выявлены 12 аллельных вариантов СА-повтора. 16-й и 17-й повторы не выявлены вообще, кстати, как и у других представителей всей индоевропейской лингвистической семьи. По частоте заметно выделяются на гистограмме 19-й, 22-й и 27-й аллели — в пределах трети от всех наличных. Самые минорные аллельные повторы (20-й и 30-й) выявлены, соответственно, в гродненской сельской группе на северо-западе Беларуси и хойникской — на юго-востоке. Равновесие Харди-Вайнберга нарушено только в последней популяции. Этот факт косвенно может свидетельствовать о наличии здесь стрессовой экологической ситуации.
В качестве критерия внутрипопуляционного разнообразия избраны показатели ожидаемой (0,83–0,87) и наблюдаемой (0,82–0,88) гетерозиготности, что позволяет считать этот маркер высокоинформативным в популяционных и этногенетических исследованиях. Беларуский этнос соответствует своей европейской характеристике по мажорным концентрациям 22-го и 27-го аллелей в сравнении с народами уральской и алтайской лингвистических семей (Лимборская С. А. и др., 2002). На территории Беларуси их присутствие одинаково возрастает в направлении с юга на север (таблица 4).
Однако 19-й аллель имеет противоположный вектор изменчивости. У минорных аллелей такая географическая зависимость не наблюдается (r = -0,59). По гетерозиготности локуса Cact685 регион ее минимальных показателей выявлен в Северной Беларуси.
Кроме проанализированных выше микросателлитных маркеров, у части беларуских популяций изучались и отдельные мини-сателлиты, в частности, высокоизменчивый локус АроВ 2-й хромосомы. Последний характеризуется значительным полиморфизмом и обладает своими собственными популяционными отличиями. Всего среди этносов Европы выявлены 26 его аллельных вариантов, в том числе у беларусов 19 (в соответствии с номенклатурой Ludwig). Таким образом, здесь отсутствуют 7 аллелей — 25-й, 33-й, 37-й, 39-й, 41-й, 47-й, 55-й. Самые распространенные — аллели с повторами 34 и 36. Уровень гетерозиготности снова высокий, он составляет от 73 до 84 процентов. Распределение частот генотипов во всех популяциях отвечает равновесию Харди-Вайнберга.
У русских тоже нет 7 аллелей, но только три из них те же, что отсутствуют и у беларусов: 41-й, 47-й, 55-й. Эти редкие аллели отсутствуют у всех трех восточнославянских этносов. Среди украинцев нет почти половины евразийских аллельных вариантов — 12 (25,27, 28, 31,33, 37, 39,41,45,47, 53, 55). Таким образом, если у беларусов вместе с русскими одинаково отсутствуют только три аллеля, то у беларусов вместе с украинцами — уже семь, или в 2,3 раза больше. Этот факт свидетельствует о большей общности исторической судьбы нашего народа с южным соседом, чем с восточным.
Итак, в Беларуси явная геногеография конкретного признака обнаружена в самом мажорном 36-м повторе аллелей: от 0,29 в Нарочанском крае, через 0,34 в Бобруйском регионе до 0,41 — в Гродненском. Почти такая же геногеография сохраняется в случае частот аллелей 30 и 48 (таблица 5).
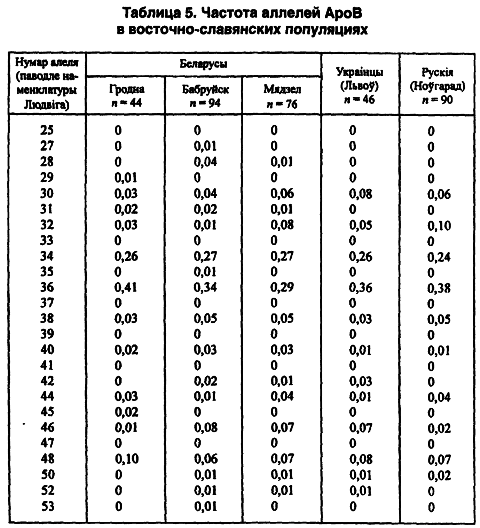
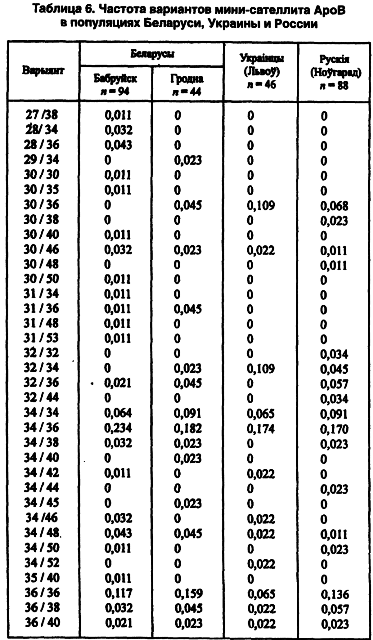
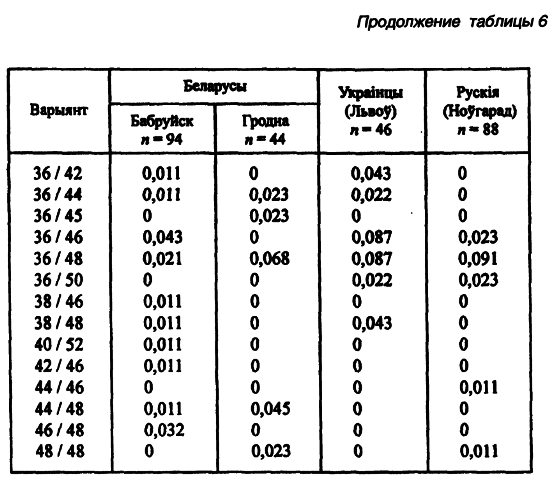
Другой модальный аллель (34-й) имеет одинаковую концентрацию во всех локальных популяциях — 0,26–0,27 (Лимборская С. А. и др., 2202). Мини-сателлит АроВ обладает выраженными расово-диагностическими свойствами. Согласно распределению его аллелей каждая из трех великих рас имеет свою особенность. Так, у европеоидов концентрация аллельных частот бимодальная, у монголоидов — унимодальная. На волжско-уральском пограничье встречаются популяции обеих этих рас. Гетерозиготность среди беларуских популяций не выходит за пределы 0,71–0,86, что полностью соответствует их европеоидности. Она не отличается от теоретически рассчитанной на статистически достоверном уровне. Все это, вместе взятое, способствовало широкому использованию данного признака в генетико-антропологических и этногенетических исследованиях (таблица 6).
Очередной маркер, полиморфизмом которого ныне широко пользуются геногеографы — мини- и микросателлитный мультилокусный ДНК-фингерпринтинг М13, называемый геномной дактилоскопией (по аналогии с классическим методом отпечатков пальцев). Большая часть вариантов минисателлита М13 отвечает критериям нормального полиморфизма. Поэтому данный метод востребован в современных популяционно-генетических исследованиях при анализе обобщенных биологических расстояний между популяциями разного иерархического уровня.
Нами по этому признаку пока изучены только популяции северо-запада и юго-востока Беларуси. С помощью компонентного и кластерного анализов в факторном пространстве действительно выявляется общее этническое «облако» (Шаброва Е. В., Микулич А. И., 2001). Анализ частотных характеристик в сравнении с финно-угорскими группами выявил значительную генетическую удаленность между ними. А по результатам многомерного шкалирования беларусы наиболее значительно отличаются от популяций коми.
Полиморфизм митохондриальной ДНК (мтДНК) в настоящее время тоже широко востребован антропогенетиками. В первую очередь потому, что она передается в наследство только по линии матери, а ее эволюция происходит за счет накопления мутаций в поколениях. Именно разнообразие митохондриального генофонда позволяет классифицировать молекулярные изменения в границах популяций и этносов. Так, если межпопуляционное различие по классическим маркерам составляет только 15 процентов от всей генетической изменчивости, то по мтДНК и генам Y-хромосомы — 30. Это весьма существенное различие, которое позволяет уточнить наши знания о геноме человека, приблизить открытие закономерностей в связях между популяциями и окружающим миром.
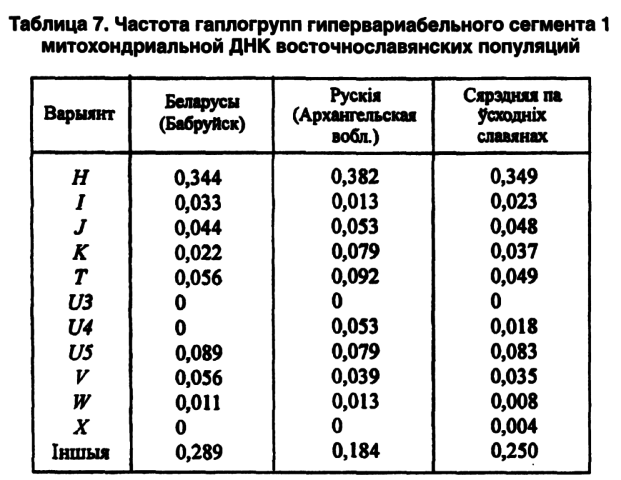
Наши совместные исследования с московскими и башкирскими коллегами показали, что индекс гаплотипового разнообразия мтДНК изменяется от 0,94 у беларусов до 0,96 у русских, 0,98 у башкиров и чувашей, 0,99 у татар и мордвы. Если учесть, что европейская раса характеризуется пониженной гетерогенностью, то наличие монгольского (или азиатского) компонента обозначается именно своей чрезвычайно высокой гетерогенностью по гаплотипам митохондриальной ДНК (таблица 7).
Среди исследованных восточнославянских популяций именно беларусы выделяются наличием гаплотипов 2, 12, 17, 22, 28. И только у нас отсутствуют 15 гаплотипов из 32 известных. По вариантам мтДНК (гаплогруппы Н, I, Т, U5, W, HV) беларусы приближаются к среднеевропейским популяциям (Лимборская С. А. и др., 2002).
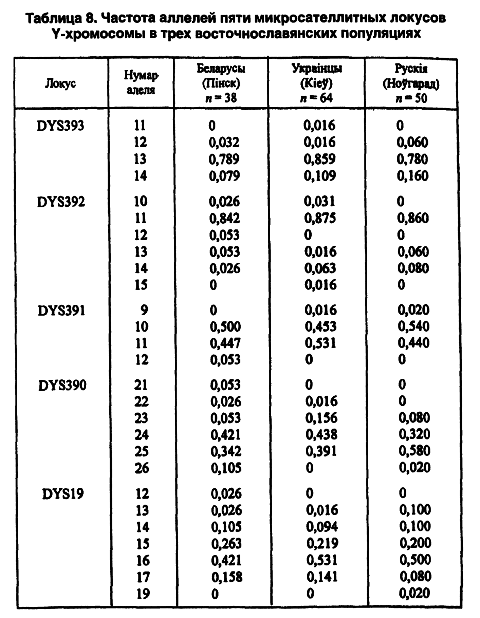
В настоящее время начато изучение полиморфных маркеров Y-хромосомы. Последняя находится в геноме мужчины и передается в наследство только по мужской линии. Y-хромосома обладает еще одной особенностью, значимой для антропологии, — чрезвычайно высокой географической и популяционной дифференциацией. Для анализа аллельного полиморфизма были выделены 5 микросателлитных локусов (DYS19, 390, 391, 392, 393) (таблица 8).
Индекс разнообразия подсчитали по формуле D = I - r2, где r — частота i-аллеля, или гаплотипа (Nei М., 1978). Достоверность различий в распределении частот аллельных вариантов и гаплотипов между обследованными популяциями определялась по Р. Фишеру (1958).