Грызуны, массами разводимые для экспериментальной работы — мыши, крысы, кролики, изучались и с точки зрения многоплодия. Все они имеют по нескольку детенышей в помете, обычно это РБ.
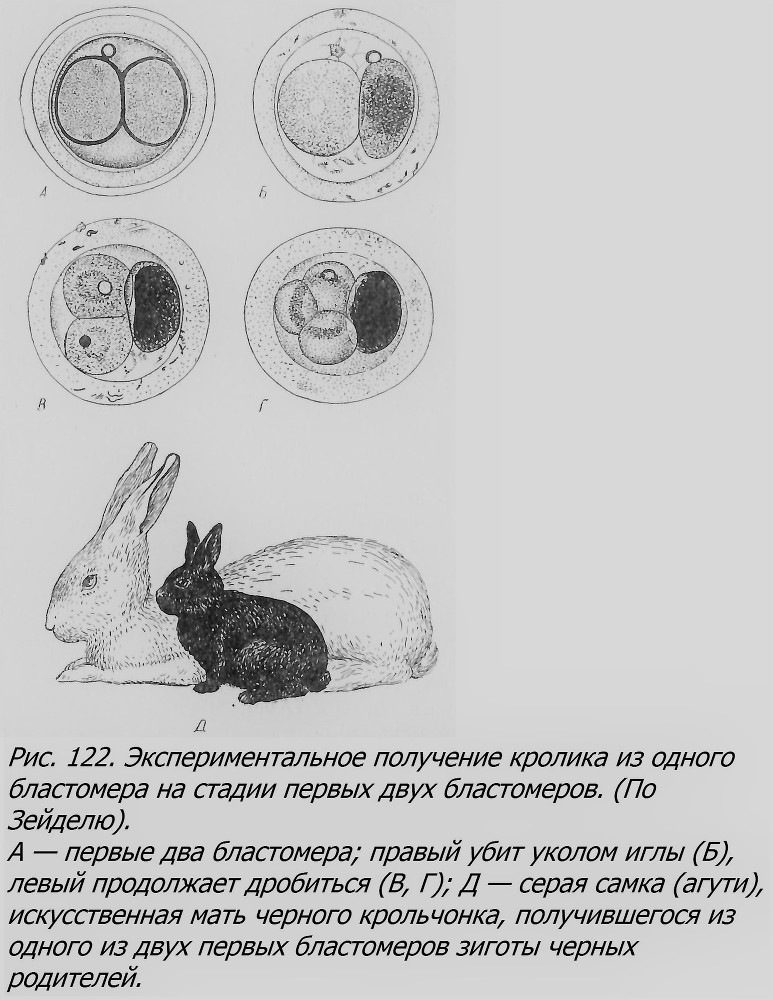
Кролики оказались удобным объектом для замечательного экспериментального показа возможности образования у них ОБ (Seidel, 1952). Извлеченные из яйцеводов зиготы кролика величиной 0.1 мм, на стадии двух первых бластомеров подвергались следующей! операции: уколом тонкой стеклянной иглы убивался один из двух бластомеров. Оставшийся бластомер мог продолжать жить и дробиться вне тела самки (рис. 122); убитый же бластомер отличался по цвету и малой прозрачности. Далее такие яйца, с продолжающим дробиться выжившим бластомером, переносились в соответственно подготовленную крольчиху. Из 27 таких «половинных» яйца два яйца закончили развитие, и у двух крольчих родилось по одному крольчонку, развившемуся из половины яйца. В пользу того, что эти крольчата именно такого происхождения, свидетельствует (кроме всех мер, которые гарантируют указанную беременность крольчих) еще тот факт, что окраска крольчат наследственно иная (они черные), чем их «матери»-инкубатора (агути). Интересно, что один из этих двух крольчат был вполне нормальный, тогда как другой имел некоторое искривление тела, напоминающее искривление тритонов, полученных Шпеманом путем перетяжки яйца (см. раздел «Амфибии»). Вероятно, искривление произошло в силу отставания развития той стороны зародыша, которая соприкасалась с убитым бластомером, повреждающе повлиявшим на эту сторону эмбриона. Предполагается, что крольчонок получился из правого бластомера.
Эти интересные факты показывают, что у млекопитающих возможно образование ОБ из первых двух бластомеров, так же как у других животных.
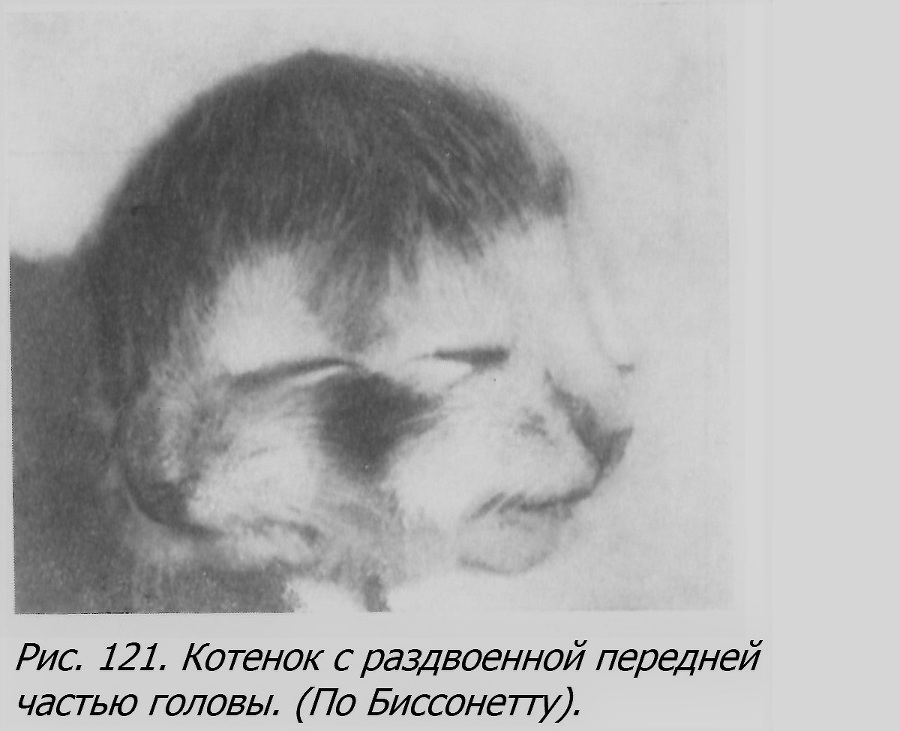
Сведений относительно ОБ у кроликов почти вовсе нет. Однако ОБ у них, по-видимому, все же возникают, так как описан случай соединенных близнецов (Fougeraux, 1786), являющийся большой редкостью. Лишь недавно описан новый случай заднего раздвоения, приблизительно с середины туловища, у кролика (d. posterior), с situs inversus у правого компонента и с некоторой разницей в форме пятна на раздвоенной части туловища (Cock, 1950).
Мыши, благодаря их плодовитости и наследственности окраски шерсти, сравнительно хорошо изученной генетически, могли бы служит удобным материалом для исследования вопросов близнечества. Но количество ОБ у мышей, по-видимому, очень мало — около 0.4—0.7% (Stewens, 1937, и др.). Описаны случаи двойных уродств. У крыс ОБ изучены очень мало.
В заключение остановимся на близнецах у обезьян, которых теперь тоже стали разводить в питомниках для опытных целей и наблюдений. Близнецы описаны у разных видов макак и других низших обезьян (Abel, 1933; Yerkes, 1934; Wislocki, 1939, и др.). У макак описан редкий случай двойного уродства — d. anterior. Животное имело две головы, двойной позвоночник, четыре руки, но один газ и одну пару нижних конечностей (Bolk, 1926). Другой случай меньшей степени переднего раздвоения описан тоже у макаки резуса (Hartman, 1943). Эти случаи свидетельствуют о существовании ОБ у обезьян, но они еще мало известны. У антропоидов (шимпанзе) описаны только РБ (Tomilin a. Yerkes, 1935, и др.). Всего до недавнего времени у обезьян установлено 24 случая близнецов. Они составляют приблизительно 1% всех родов, как у человека. Большинство близнецов — РБ (Schultz, 1956).
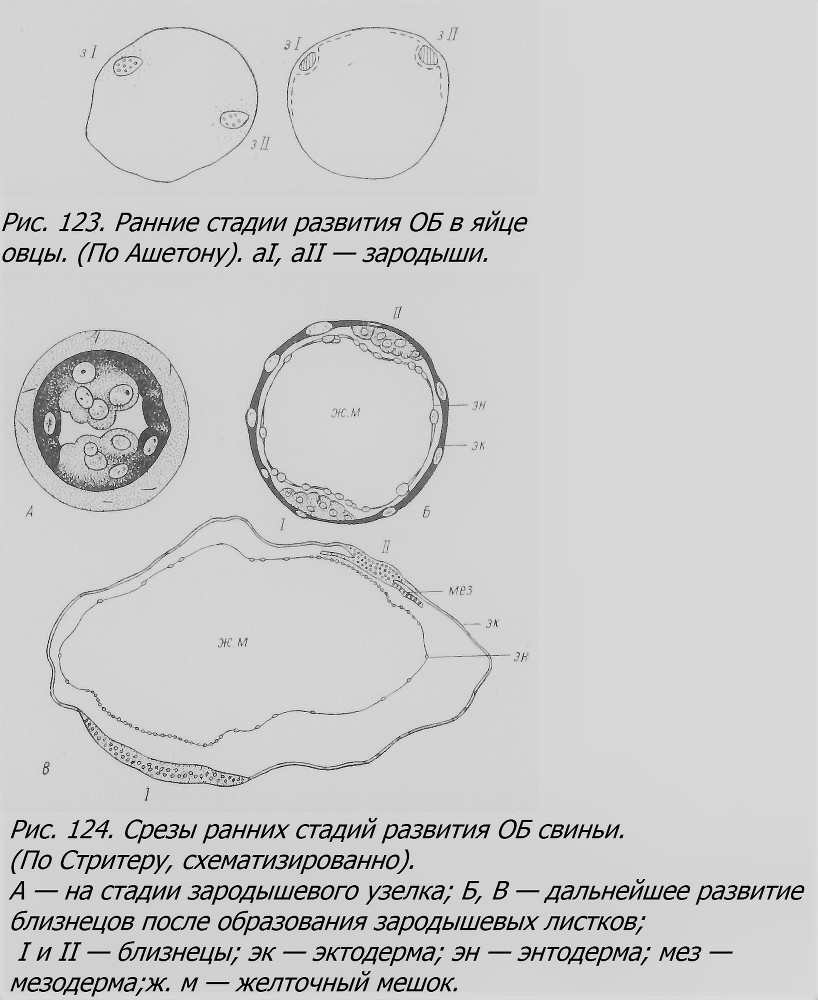
Эмбриогенез близнецов, особенно ранний, у рассмотренных млекопитающих плохо изучен, так как материал очень редок. У овцы описана только одна бластоциста в возрасте около 7 дней с двумя зародышевыми узелками в период образования эндодермы (рис. 123), т. е. образования первичной полоски (Assheton, 1898, 1913). Другой случай ранней стадии развития ОБ в возрасте около 13 дней был описан у свиньи (Streeter, 1924). В матке одной свиньи среди нормальных яиц было обнаружено одно яйцо в стадии имплантации. Гистологическое изучение этого яйца показало, что оно уже прошло стадию бластоцисты и имело два расположенных друг против друга зародышевых диска с одним общим желточным мешком. Энтодерма была больше развита в области соприкосновения с каждым из зародышевых дисков (рис. 124). Один из зародышей (I) несколько отставал в развитии от другого (II): у последнего образование мезодермы было более выражено, чем у первого. Оба зародыша, по-видимому, находились на ранней стадии первичной полоски, т. е. в начале гаструляции. Стритер предполагает, что раздвоение яйца произошло во время образования бластоцисты: возникло два зародышевых узелка вместо одного. Чем это вызвано, остается неясным. У свиньи установлено вообще много аномалий раннего развития зиготы: около 10% яиц вообще не дробится, около 10% дегенерирует в стадии бластоцисты, от 5 до 10% развивается ненормально, и только около 70% превращается в жизнеспособных поросят. ОБ и в зародышевом состоянии у свиньи обнаруживаются редко. У других млекопитающих ранние стадии эмбриогенеза почти неизвестны. У облученных х-лучами мышей среди 1500 эмбрионов найден один случай ОБ, с общим желточным мешком, на стадии первичной полоски. Один из близнецов несколько более развит, чем другой. Оба были приблизительно на той же стадии развития, как и смежные одиночные эмбрионы. По-видимому, никакой задержки развития зиготы, из которой получились эти близнецы, не было; это противоречит гипотезе Ньюмена (Bodemann, 1935).
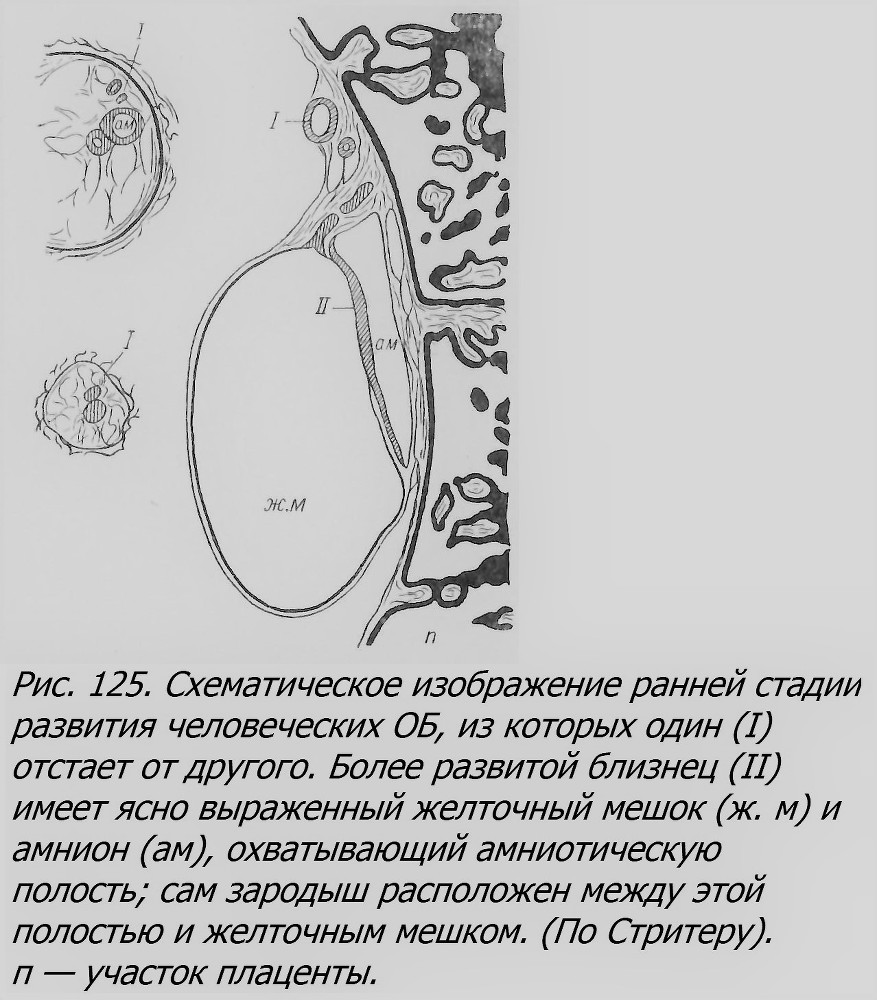
Мало материала имеется в распоряжении науки также относительно ранних стадий развития ОБ у человека. До недавнего времени был известен только один препарат на стадии первичной полоски (рис. 125), описанный Стритером (Streeter, 1919). Один из близнецов заметно опередил своего партнера, имеет амнион и желточный мешок. Длина этого зародыша 0.92 мм. Другой заметно меньше. Близнецы возникли, как предполагает Стритер, на стадии бластоцисты — образовалось два зародышевых узелка, подобно тому, как это показано на схеме Паттена (рис. 126).
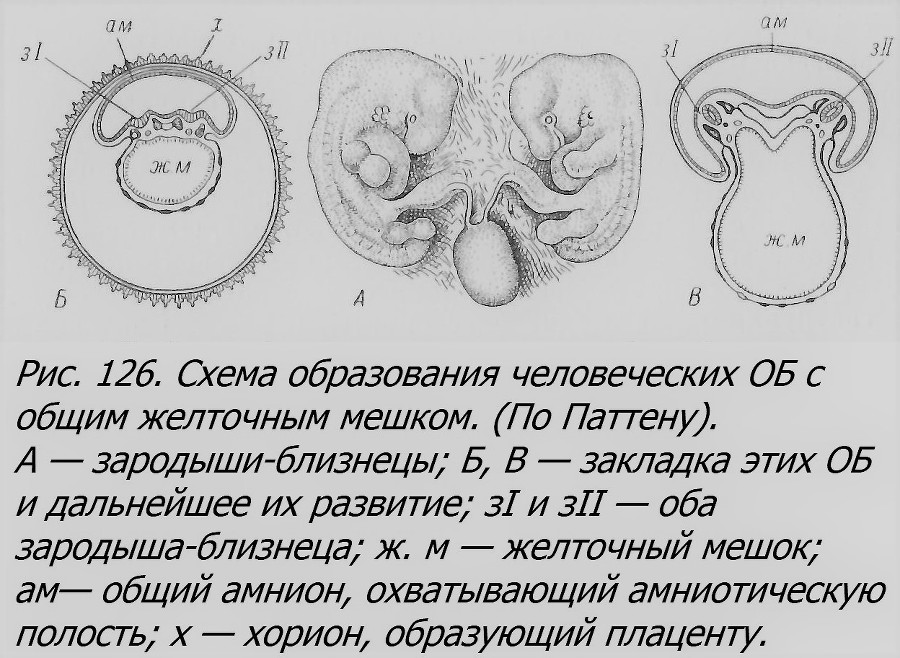
Недавно описан еще один препарат человеческих ОБ на стадии первичной полоски, в возрасте около 17 дней (Corner et Baltimore, 1955). Эта пара считается самой юной из всех известных. Эти близнецы имеют общий хорион, но разные амнионы и желточные мешки. Другая пара, недавно описанная Мортоном (Morton, 1954), обнаруживает начальные стадии развития головы и конечностей. Этим зародышам близнецов около 32 дней. Они, как и предыдущая пара, имеют общий хорион и разные желточные мешки и амнионы (Corner et Baltimore, 1955). Из немногочисленных пар ОБ в возрасте 5—6 недель отметим еще пару монохориальных зародышей (рис. 126), одинаковой величины, длиной около 12 мм, в возрасте около 6 недель с общим желточным мешком (Агеу, 1922а). Возможно, что они возникли также на стадии бластоцисты, но можно предположить, что они произошли в результате двойной гаструляции (Newman, 1923).
Но вопросу об эмбриогенезе близнецов и, в частности, ОБ у человека на ранних стадиях развития эмбриона мы уже говорили в связи с вопросом о зародышевых оболочках и причинах возникновения близнецов (см. главу четвертую).
Весь рассмотренный небольшой материал по эмбриогенезу ОБ у млекопитающих, от броненосца до человека, показывает, что и у этого класса, как и у других животных, ОБ, очевидно, возникают тоже только на ранних стадиях развития: может быть, еще при первом дроблении зиготы, вероятно чаще всего до гаструляции, на стадии бластоцисты, и реже — на ранних этапах гаструляции.
Глава девятая
КАК ЧАСТО РОЖДАЮТСЯ ЧЕЛОВЕЧЕСКИЕ БЛИЗНЕЦЫ?
Для многих видов животных многоплодие оказывается нормой, обычным явлением при размножении. Не пускаясь в рассмотрение всего многообразия этих феноменов, отметим лишь, что у различных видов млекопитающих самка, как правило, производит на свет несколько детенышей - обычно РБ, например у мышей, крыс, кроликов, кошек, собак, свиней и т. д. Рождение одного детеныша у таких животных исключение. Наоборот, для других видов это правило, а многоплодие исключение; сюда относятся корова, лошадь, слон и другие. До некоторой степени промежуточное положение в этом отношении занимают овцы, козы и другие животные. Для некоторых видов многоплодие, по-видимому, установившийся видовой признак, так же как для других таковым является одноплодие. И есть виды, у которых это свойство нечетко выражено, меняется в зависимости от породы, линии и т. д. О связи многоплодия с размерами тела вида или расы речь была выше (см. главу третью).