Изучая процесс гаструляции в курином яйце с помощью стереокинематографии Грэпер (Graper, 1931) наблюдал своеобразное движение тканей во время инвагинации, т. е. перемещения тканей в бластопор при образовании внутреннего слоя зародыша — энтодермы. Схематизируя возможные комбинации тканевых потоков при двойной гаструляции, в зависимость от которой Грэпер ставит возникновение близнецов, он старался «единообразно» объяснить образование различных типов соединенных близнецов (см. систему Швальбе, стр. 73). Схема Грэпера в наше время, однако, утратила свою универсальность и может годиться только для объяснения некоторых случаев (ср. Politzer, 1955).
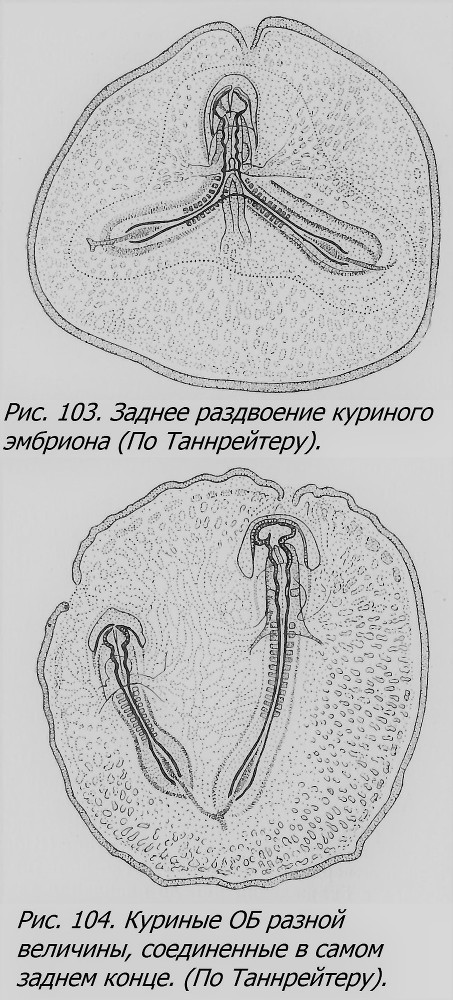
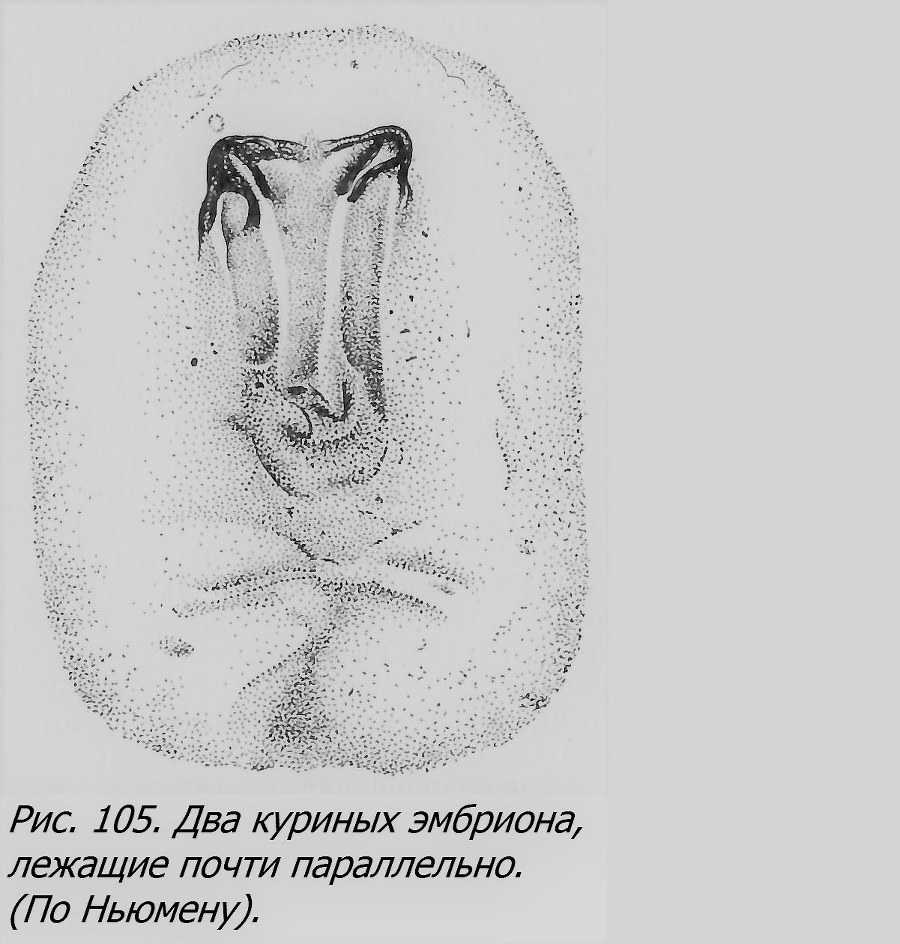
Чем вызвано образование близнецов у птиц? Еще Дарест (Dareste, 1891), а позже Стокард (Stockard, 1921) и Ньюмен (Newman, 1923) высказали предположение, что близнецы возникают в тех яйцах, которые оказываются снесенными до окончания «критического» периода, когда еще возможно образование близнецов. Предполагается, что снесенное яйцо подвергается резкому охлаждению, которое вызывает задержку развития на некоторое время — это и способствует возникновению двух пунктов развития в силу их физиологической изоляции. Это то же самое объяснение, с которым мы уже встречались, касаясь образования близнецов у морских звезд, рыб, лягушек и т. д. Если допустить, что охлаждение застает яйцо на сравнительно поздней стадии развития — гаструляции, то вероятнее всего ожидать, что один, уже образовавшийся эмбрион подвергается расщеплению, т. е. близнецы чаще всего будут возникать по третьему способу, что, по мнению Ньюмена, и имеет место, притом с раздвоением заднего конца чаще, чем переднего, так как задний конец формируется позже переднего. В связи с этой гипотезой были поставлены опыты охлаждения яиц голубей, извлеченных досрочно из тела птицы (Riddle, 1923). Эти опыты не подтвердили гипотезы. Другая попытка проверки ее заключалась в охлаждении яиц до окончания гаструляции путем охлаждения самой птицы (курицы) в период начального развития в ней оплодотворенных яиц, до их кладки (Sturkie, 1946). На 73 обследованных так охлажденных яйцах было обнаружено 6 двоен, т. е. 8.2%, тогда как в норме двоен бывает около 0.111% (Byerly a. Olsen, 1934), т. e. увеличение близнецов на 8.09%, цифра статистически достоверная. Все экспериментальные эмбрионы отставали в развитии, из них 32.9% имели аномалии развития, не считая раздвоения. Близнецы были разных типов, все соединенные; малое число двоен не дает возможности сколько-нибудь основательно судить о численном отношении типов двойных уродств. Однако этими данными нельзя считать гипотезу Стокарда—Ньюмена доказанной, нужны дальнейшие опыты для ее проверки.
Близнецов у птиц вызывали и другими экспериментальными методами. Пользуясь методом Шпемана, трансплантировали кусочки различных тканей, могущих индуцировать образование в бластодерме добавочного зародыша — искусственного близнеца (Morita, 1936; Waddington, 1941). Эти явления аналогичны описанным у амфибий.
Путем механической изоляции — поперечным разрезом скальпелем — можно вызвать образование близнецов у утиного эмбриона (Lutz, 1953). Ориентация головы переднего партнера зависит от стадии гаструляции. Если разрез пройдет через участок, куда уже проникла образующаяся энтодерма (вспомним, что этот процесс идет от заднего конца будущего зародыша к переднему), то голова переднего близнеца будет ориентирована нормально, как и голова заднего, т. е. в одном направлении. Однако голова переднего эмбриона окажется ориентированной назад, навстречу голове заднего, если разрез провести на той стадии, когда энтодерма еще не образовалась в переднем участке. Это Лутц объясняет тем, что в этом последнем случае образование энтодермы потечет от переднего края бороздки, т. е. в обратном направлении, и потому голова переднего близнеца образуется у линии разреза, иначе говоря, будет обращена к голове заднего близнеца. Таким образом можно произвольно изменять относительное направление осей обоих близнецов.
Близнецов у птиц можно получить еще и другим приемом. На стадии ранней гаструляции, пока первичная полоска составляет одну четверть светлого поля бластодиска (area pellucida), наносится повреждение с помощью ультрафиолетового укола (по методу Чахотина) или электротока. В результате может возникнуть раздвоение процесса гаструляции и образование двоен, а изредка — троен. После того как первичная полоска достигла половины своей длины, близнецы этим методом уже не получаются (Twisselmann, 1939). Различные варианты получаемых близнецов зависели от конкретных условий опытов.
Таким образом, и стадия эмбриогенеза — не позже гаструляции, и приемы, вызывающие экспериментальное образование ОБ у птиц, в общем, аналогичны таковым у амфибий. Это говорит о сходстве по существу тех процессов, которые лежат в основе полиэмбрионии у этих животных.
Млекопитающие
Среди этих животных есть два вида из отряда неполнозубых: броненосцы-армадилы (Dasypus novemcinctus и D. hybridus), которые замечательны тем, что закономерно размножаются путем образования нескольких зародышей из одного яйца (пол близнецов одного помета всегда одинаковый). Первый вид обычно имеет четырех однояйцевых близнецов, второй — разное число (до 12). Это явление полиэмбрионии у млекопитающих в известной мере аналогично вышеописанным случаям у кольчатых червей и паразитических насекомых. У млекопитающих это очень редкое явление. Другие виды броненосцев размножаются обычным путем, ОБ у них не описаны. У многих видов млекопитающих ОБ явление далеко не постоянное, наоборот, как мы увидим из дальнейшего, ОБ у остальных видов млекопитающих скорее исключение, чем правило.
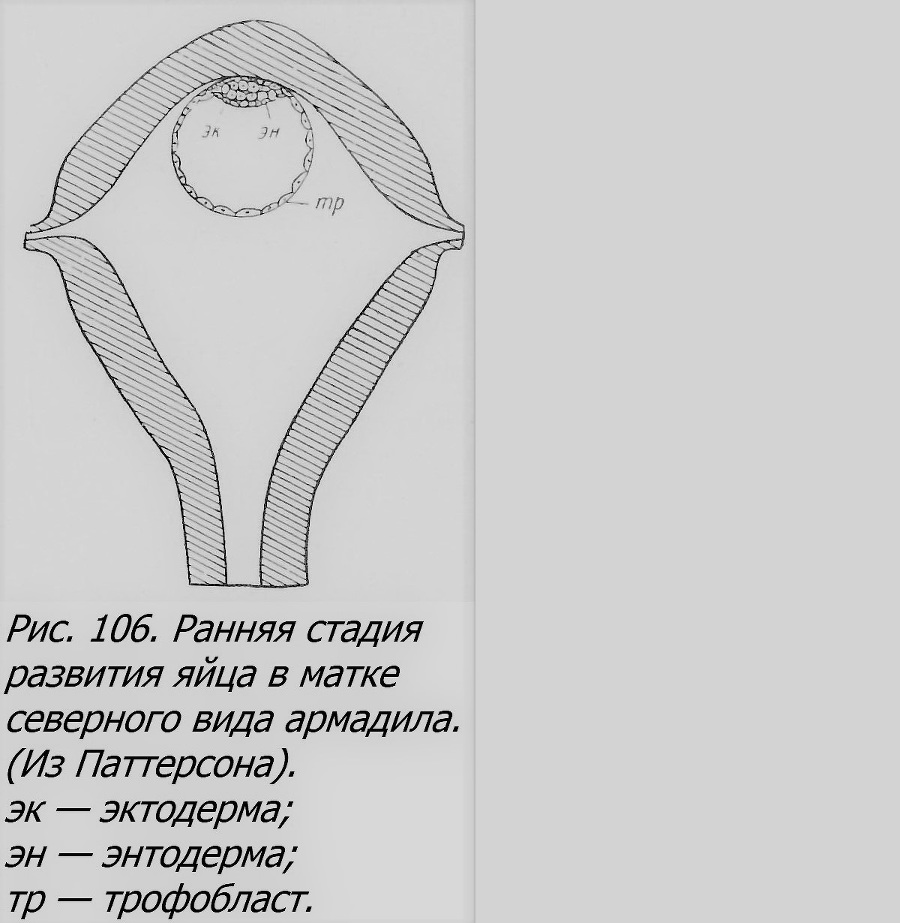
Оба вида броненосцев, имеющие ОБ, живут в Америке. Их удивительный способ размножения был давно известен. В науке он впервые описан в 1885 г. Иерингом, а подробно изучен только к начале XX в. Фернандец (Fernandez, 1909, 1913) изучил образование ОБ у южной формы (D. hybridus), а Ньюмен и Паттерсон (Newman а.Patterson, 1909; Patterson, 1913, 1927; Newman, 1923) — у техасского броненосца (D. novemcinctus). Этот последний был изучен подробнее, и потому мы его и рассмотрим в качестве примера. В пользу однояйцевости его близнецов прежде всего говорит тот факт, что возникновение их сопряжено с образованием в яичнике только одного желтого тела (Newman, 1923), что обычно связано с выходом одного яйца из фолликула, который и превращается в желтое тело яичника. Развитие яйца прослежено от поздних стадий дробления до полного развития близнецов (Patterson, 1913). В период половой активности, в августе, из оплодотворенного яйца начинает развиваться зародыш. В яйцеводах самки в это время можно найти самые ранние стадии эмбриогенеза, до бластоцисты включительно, т. е. до стадии полой сферы, ограниченной слоем клеток (трофобластом) с утолщением в одном участке — зародышевым узелком, из которого возникает сам зародыш (рис. 106).
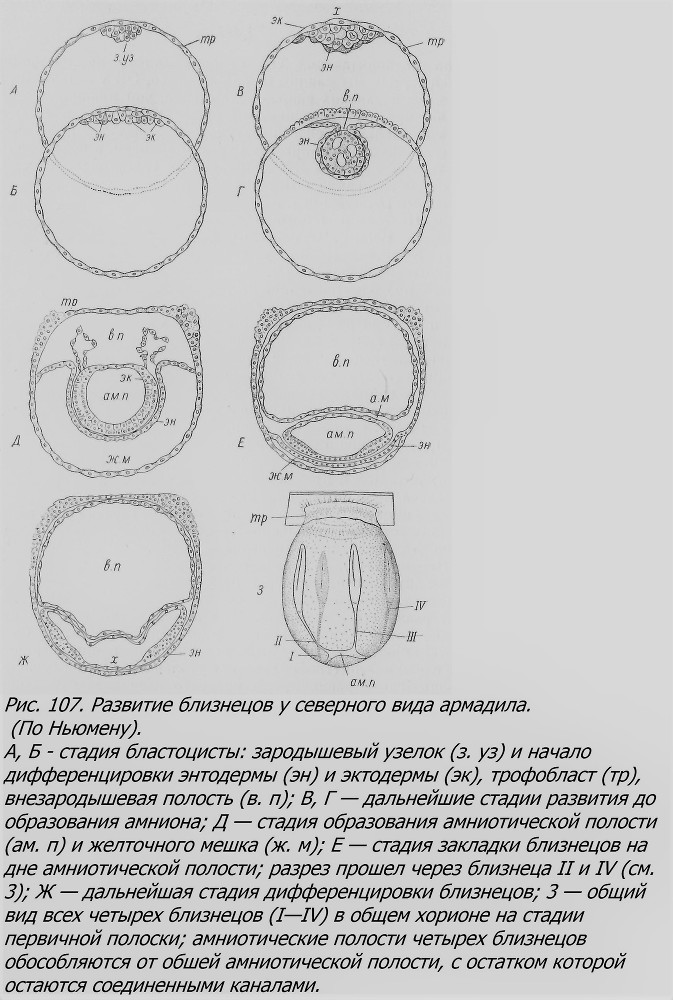
Бластоцнста, попав в матку, не сразу внедряется в ее стенку, а несколько недель остается неимплантированной. Это «период покоя», когда развитие, по-видимому, почти полностью останавливается. Осенью, после прикрепления к стенке матки, развитие бластоцисты возобновляется, при этом часть стенки бластоцисты превращается в плаценту, а в зародышевом узелке дифференцируются экто- и энтодерма (рис. 107). В разросшейся эктодерме развивается полость — это амниотическая полость, над ней возникает другая полость — внеамниотическая, и под амниотической полостью — желточный мешок; верхнюю стенку его образует энтодерма. На дне амниотической полости эктодерма утолщается — из нее в дальнейшем образуется зародышевый диск; это тот участок, где возникает сам зародыш. Полость амниона сплющивается и образует два боковых выроста, которые в дальнейшем в свою очередь раздваиваются. Так получается центральный участок амниотической полости и четыре ветви ее (рис. 108). В каждой из них затем возникает по первичной полоске, т. е. закладывается ось зародыша. Все оси, как лучи, обращены к общему центру, головным концом к нему (рис. 107, 3). В дальнейшем все четыре зародыша вытягиваются в длину, головы их обращены в противоположную от плаценты сторону. Таким образом, появлению зародышей близнецов предшествует своеобразное «почкование» амниотической полости с повторным удвоением. Несколько иначе протекает процесс разветвления амниотического пузыря у южного вида броненосца, где выросты («почки») возникают как бы беспорядочно, в разной последовательности, и оказываются различного размера и неодинаково расположенными по отношению друг к другу (рис. 109).