Таким образом, по отношению к соединенным ОБ гипотеза Ньюмена кажется несколько более обоснованной, чем по отношению к свободным ОБ. Однако достаточно доказанной ее все же считать нельзя, и для полного выяснения правильности её или степени правильности необходимы дальнейшие исследования. Во всяком случае, в качестве рабочей гипотезы она была полезна и остается таковой и в настоящее время. Многие вопросы эта гипотеза до сих пор не может объяснить. Сам Ньюмен (Newman, 1940b) указал некоторые из них. Почему среди ОБ могут чаще встречаться признаки зеркальности, чем среди одиночек? Почему у ОБ одни признаки оказываются зеркальными, а другие нет — например при s. i. у одного из близнецов теменной завиток у него идет нормально? Почему у одних свободных ОБ с одинаковой степенью сходства зеркальность выступает резче, а у других слабее? Почему у одинаково соединенных близнецов может быть разное выражение зеркальности? Нет надобности увеличивать число таких вопросов; приведенных достаточно, чтобы показать, насколько вопрос о зеркальности у близнецов еще неясен, как много еще непонятного в нем.
Другого рода гипотезы были высказаны для объяснения зеркальности асимметричных признаков на основе генетики. В области наследственности асимметрии есть несколько интересных исследований, например Пшибрама (Phzibram, 1908) о наследственности разного цвета правого и левого глаза у ангорских кошек или Бойкотта с сотрудниками (Boycott et al., 1929) над улитками, имеющими спирально завитую раковину направо или налево. Объяснение, даваемое этим явлениям, сводилось на менделирование определенных генов. Дальберг (Dahlberg, 1926, 1929, 1943/44) по аналогии с этими явлениями высказал предположение о существовании неодинакового распределения генов при образовании ОБ, отчего зависит зеркальность асимметричных признаков, назвав свою гипотезу «теорией генотипической асимметрии». Эта гипотеза носит умозрительный характер и недостаточно подтверждается фактами. Несколько иначе, но в том же направлении Боутервек (Bouterwek, 1943) старался доказать, что асимметрия и зеркальность ОБ основаны на различии их генотипов. Этот тезис и его доказательства вызвали ряд возражений (Lotze, 1937, и др.), и действительно, попытки Боутервека не производят убедительного впечатления. Вопрос о значении генотипа для зеркальности у близнецов остается пока что нерешенным. Генотип, вероятно, играет известную роль и, может быть, неодинаковую у разных групп животных (Torgersen, 1950). Однако понять роль генотипа можно будет только тогда, когда глубже будет изучен физиологический механизм становления асимметрии зародыша, изменение этой асимметрии в зависимости от раздвоения зародыша и в связи с этим возможности возникновения зеркальности на разных ступенях развития эмбриона. Тогда будет также изжит односторонний характер гипотезы Ньюмена.
Теперь, после рассмотрения некоторых общих вопросов близнецовой проблемы, мы можем перейти к знакомству с многообразным проявлением близнечества в различных классах животного царства, начиная с низших форм и кончая человеком.
Глава восьмая
БЛИЗНЕЦЫ У ЖИВОТНЫХ
Беспозвоночные
Мы начнем рассмотрение образования близнецов в этой обширной группе с представителей низшего типа многоклеточных животных — кишечнополостных.
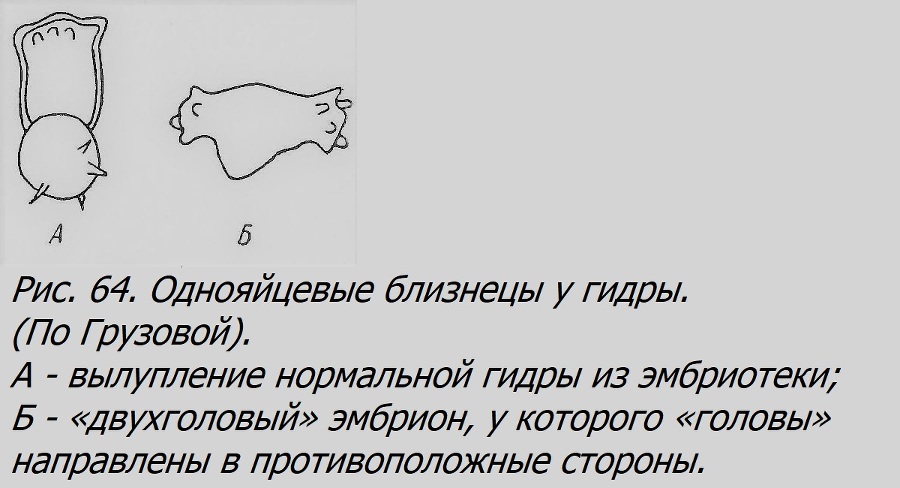
У гидры ОБ до последнего времени не были описаны. Совсем недавно установлены единичные случаи вылупления из эмбриотеки «двухголовых» эмбрионов, которых, очевидно, надо считать ОБ (рис. 64; Грузова, 1956).
До известной степени аналогичное явление установлено на почках гидры, причем особи, возникшие из «близнецовых» почек, в свою очередь образовывал и такие почки приблизительно в два раза чаще, чем особи, возникшие из обыкновенных почек (Turner, 1956).Конечно, о почках-«близнецах» можно говорить только с известным насилием над, термином «близнецы». Вообще же вопрос о близнецах у гидр еще недостаточно выяснен и должен быть обстоятельно изучен. Но у некоторых других форм кишечнополостных ОБ получены экспериментально. Так, если начавшую дробиться зиготу (оплодотворенное яйцо) гидроида клитии (Clytia flavidula) разрезать тонкой острой иглой пополам на стадии первых двух клеток (бластомеров), т. е. полностью изолировать их друг от друга, то каждый из этих двух бластомеров будет продолжать дробиться, сначала как бы в контакте с другим, а потом, закругляясь, образует самостоятельный комплекс клеток. В процессе дальнейшего развития получается бластула[9], гаструла и, наконец, молодой полип, но только половинного против нормы размера (рис. 65).
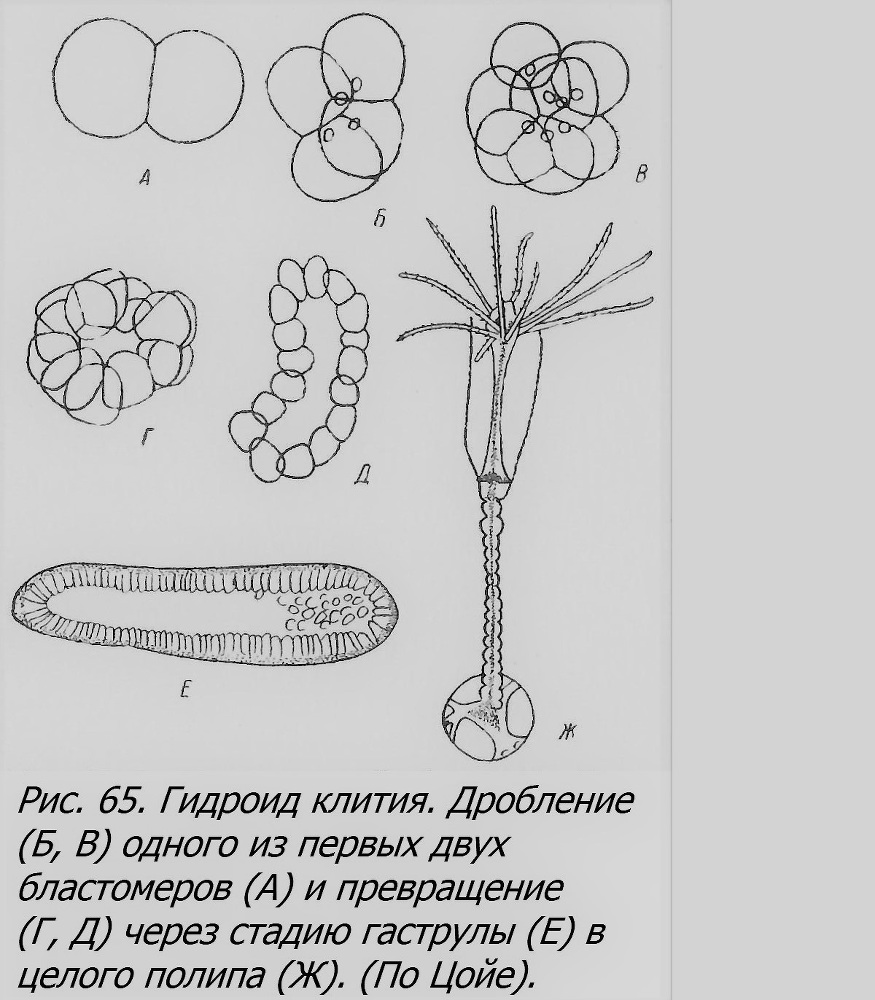
Если таким же способом изолировать первые четыре бластомера, то развитие пойдет аналогично, и получится четыре целых полипа, соответственно меньшего размера. И число щупалец у них вначале будет меньше обычного. Но уже при изоляции на стадии восьми бластомеров развитие каждого бластомера не идет и дальше гаструлы. Похожие результаты получены и на других гидроидах (Zoja, 1895), а также на высших кишечнополостных — сцифомедузах. Недавно колонии-близнецы получены у гидроидного полипа Hydractinia путем разделения первых двух бластомеров (Hauenschild, 1954).
Известно, что гидроиды и, в частности, пресноводные гидры в большой мере способны из небольших кусочков тела давать целое животное. Это явление регуляции можно до известной степени аналогизировать, с регуляцией отдельных бластомеров.
Яйца кишечнополостных нежны и легко разрушаются, с ними трудно ставить опыты и по ряду других причин.
Более удобным объектом для экспериментального получения близнецов являются иглокожие: морские ежи и морские звезды. Как и у гидроидов, близнецы у морских ежей получены путем изоляции бластомеров на самых ранних стадиях дробления яйца, стадиях двух и четырех бластомеров. На более поздних стадиях изолированные бластомеры уже не способны дать целый организм.
Изоляция достигается разными методами. Самый простой и грубый — это встряхивание сосуда, содержащего воду с дробящимися яйцами морского ежа (Driesch, 1892). Лёб (Loeb, 1894) помещал яйца морского ежа через 10 минут после оплодотворения в морскую воду, наполовину разведенную пресной водой. Яйца разбухали и лопались. Часть плазмы вытекала и закруглялась, не теряя связи с яйцом. После перенесения яиц в нормальную морскую воду развитие продолжалось; вытекшая плазма получала в процессе дробления ядро, и из нее тоже развивался зародыш, т. е. возникала пара близнецов, иногда тройня или четверня. Изящнее способ Гербста (Herbst, 1900): в искусственной морской воде, лишенной солей кальция, слой эктоплазмы растворяется, и бластомеры легко отделяются друг от друга, после чего их возвращают в нормальную морскую воду. Этот способ был развит и видоизменен Лёбом (Loeb, 1909), получавшим до 90% ОБ, частью симметричных (рис. 66),
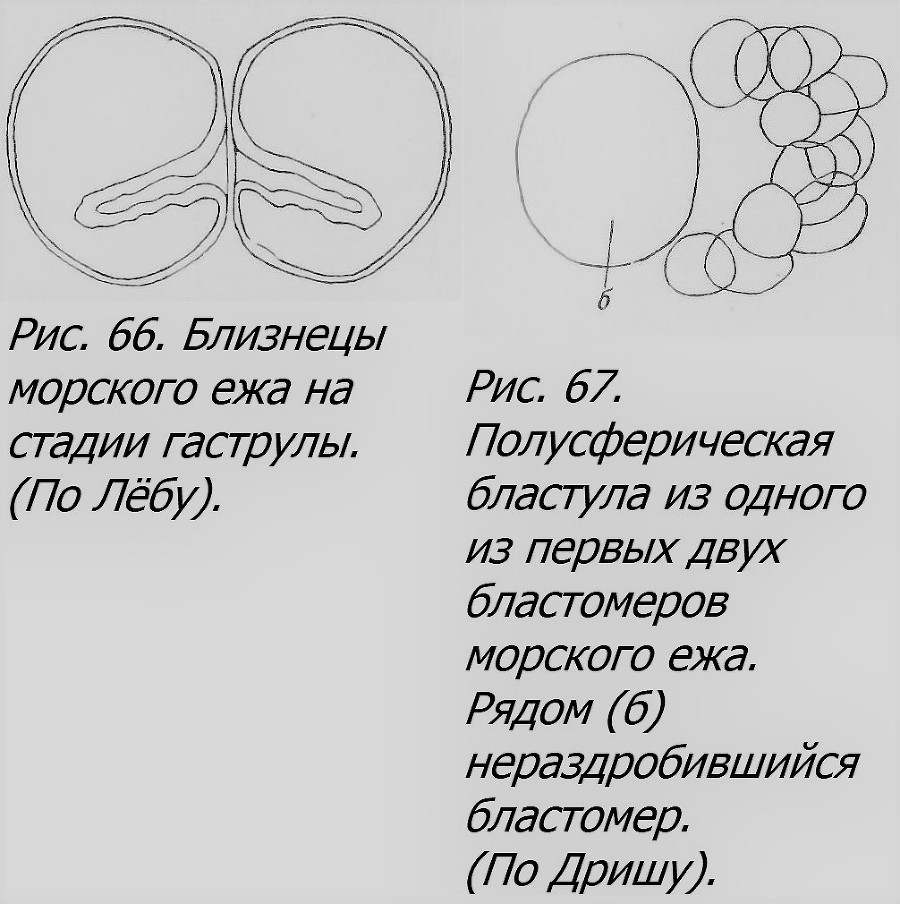
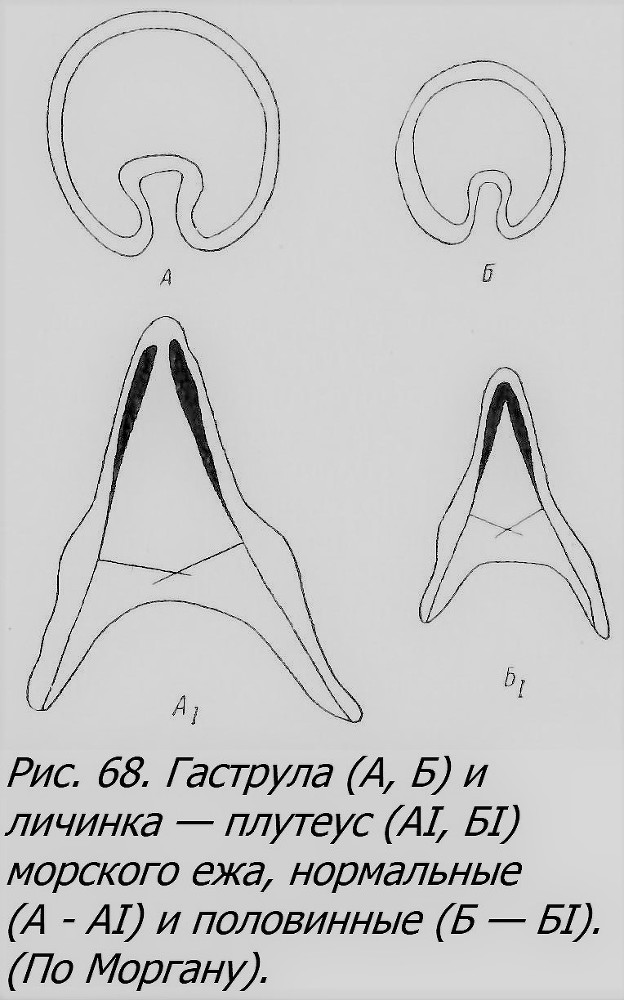
так как они возникали из первой пары бластомеров, физиологически изолированных. Существуют и другие способы (Harvey, 1940). Интересно отметить, что у некоторых видов морских ежен каждый из изолированных первых двух бластомеров, дробясь, образует полусферическую бластулу, «открытую», как при нормальном дроблении, когда рядом с ней другую полусферу образует парный с ней бластомер (рис. 67). Лишь при дальнейшем развитии бластула, замыкаясь, приобретает сферическую форму, а затем переходит в гаструлу и далее в личинку (плутеус), нормальной формы, но половинного размера (рис. 68).
Близнецы могут некоторое время развиваться, будучи слабосоединенными слизистой массой наружной поверхности их тел; но, начав свободно плавать, бластулы-двойни скоро разъединяются. Развитие близнецов у морских ежей может идти с разной скоростью, иногда с возникновениями разных аномалий. Как правило, по-видимому, развитие протекает тем медленнее, чем меньше размеры зародыша, в связи с тем, что он образовался из половины яйца (Horstadius, 1928 ). Аналогичное явление обнаружено у ланцетника (Morgan, 1927), тритона (Spemann u. Falkenberg, 1919) и у других обектов. Замедление развития наблюдается и у нормальных особей и, по-видимому, зависит преимущественно от различных внешних влияний на ход развития. Нет никаких данных для допущения, что между первыми двумя бластомерами было бы какое-либо существенное различие, что они, например, уже предназначены стать правой и левой половиной будущего зародыша. То же можно сказать и относительно первых четырех бластомеров — они, по-видимому, равноценны; это видно из того, что из них получаются целые зародыши-четверни.