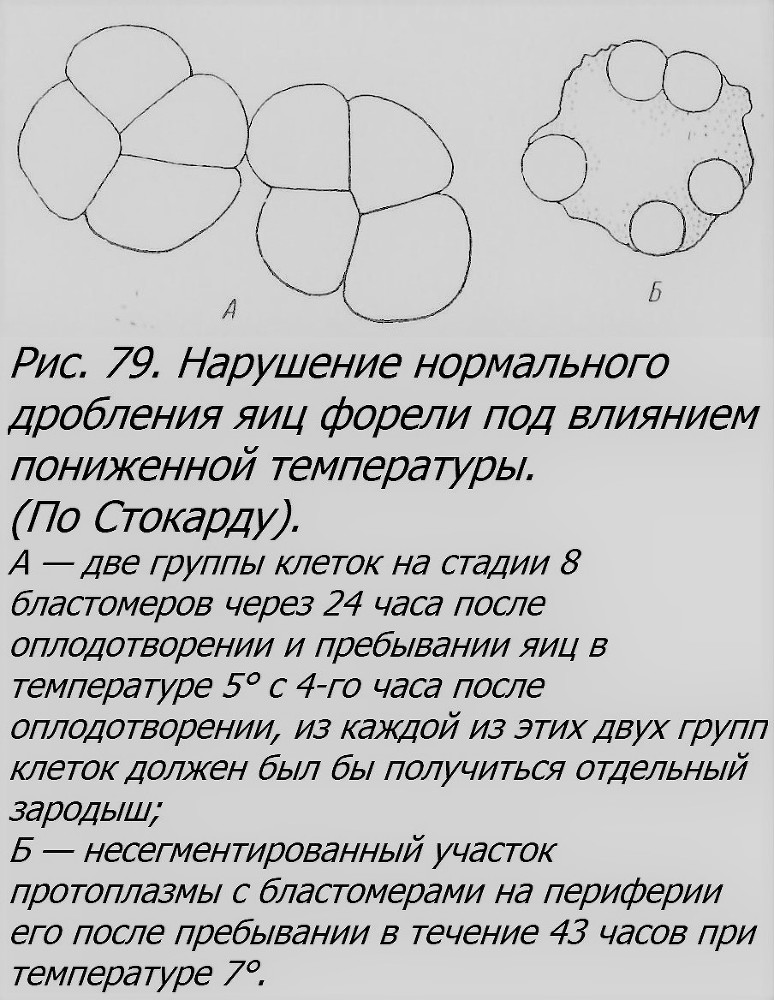

Гораздо лучше исследованы близнецы у высших рыб, костистых, как найденные в природе, где они в общем довольно редко встречаются, так и полученные в эксперименте. ОБ у костистых рыб могут возникать, по-видимому, по крайней мере двумя путями: 1) путем образования добавочных краевых узелков, т. е. пунктов, откуда начинается гаструляция, и 2) путем расщепления надвое по продольной оси первоначально одного эмбриона. Первый путь, по-видимому, имеет несколько вариантов. Могут оказаться физиологически изолированными группы бластомеров (рис. 79), которые, возможно, дадут начало разным зародышевым дискам. Могут возникнуть разные точки гаструляция, в разной мере удаленные друг от друга на бластодиске (рис. 80). Если они закладываются на противоположных краях диска и растут навстречу друг другу, то получатся более или менее нормальные зародыши, соединенные только общим желточным мешком или тканями тела в области брюха (рис. 81). Не всегда партнеры оказываются равновеликими и равноценными. Наблюдаются различные ступени перехода к комбинациям, когда один из близнецов оказывается развитым относительно нормально, а другой — отставшим в развитии, с дефектами в формировании различных частей тела (головы, глаз, туловища, конечностей и т. д.), словом, здесь создаются отношения типа автозит-паразит (рис. 81, Б — Г). Близнецы рыб, соединенные в брюшной области, морфологически аналогичны близнецам млекопитающих и человека, которые называются торакопаги, и близким к ним формам.
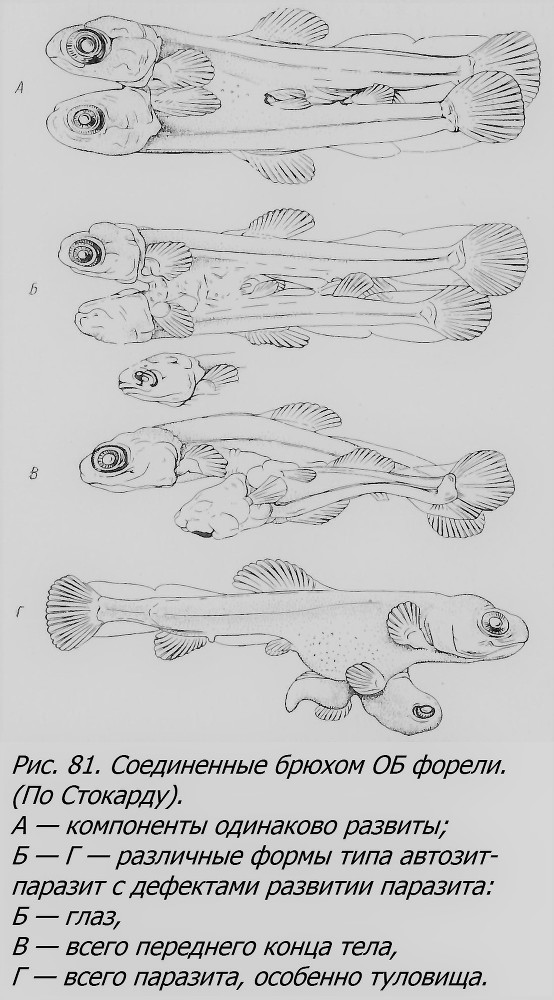
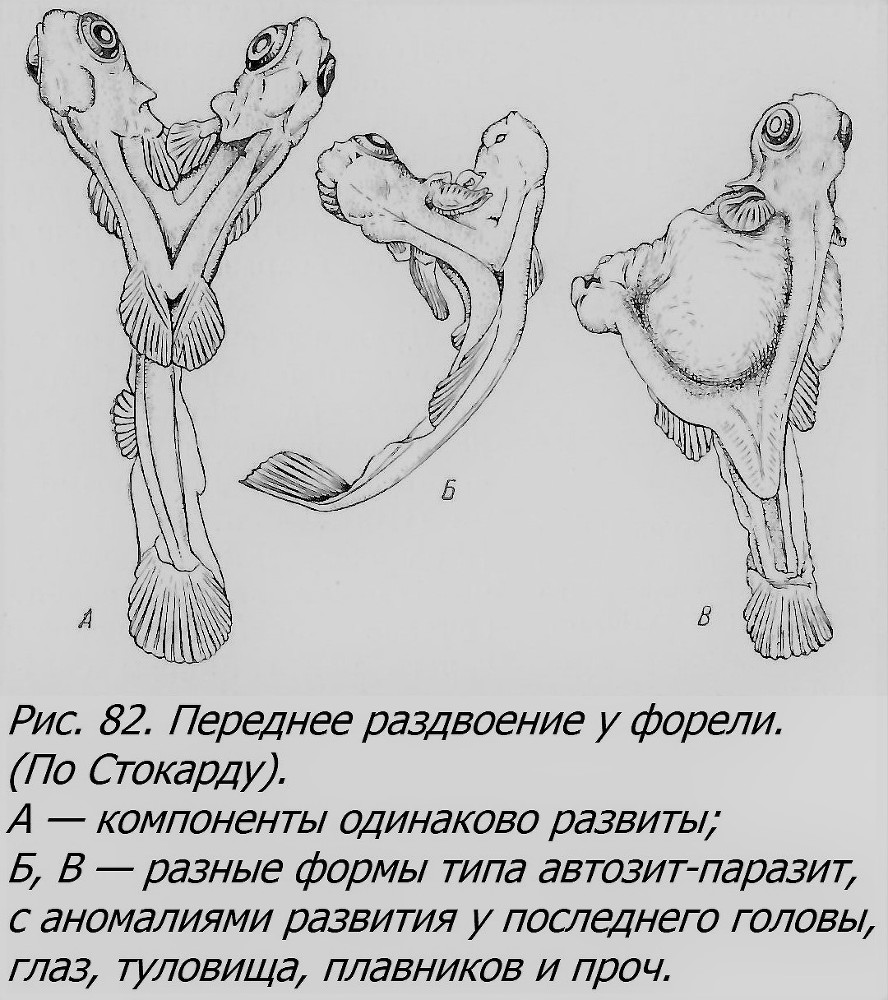
В случае, если закладка двух и более близнецов произойдет на диске по соседству друг от друга, то в процессе роста и развития зародыши могут сблизиться и в той или иной мере срастись, образуя как разные формы равновеликих соединенных близнецов — симметричных (рис. 82), так и неодинаково развитых, где тоже может выступить отношение автозит-паразит в разной степени контраста. В отличие от первого типа партнеры будут здесь соединены боками, чаще имея две разные головы, чем два хвоста; например, до 90% всех таких двойных уродств форели имеют две головы (Lynn, 1938). Это тип, уже нам встречавшийся, называемый duplicitas anterior, т. е. переднее раздвоение, в отличие от d. posterior — заднего раздвоения. Переднее раздвоение может быть различной степени: от едва заметного снаружи раздвоения головы при полной норме всего тела (рис. 83) до выраженного в той или иной мере раздвоения не только головы, по и переднего участка тела (рис. 84). Интересно, что в таких случаях у одного из партнеров, чаще правого, нередко внутренние органы лежат в порядке, обратном норме, зеркально отражая обычное расположение внутренностей, т. е. имеется situs inversus.
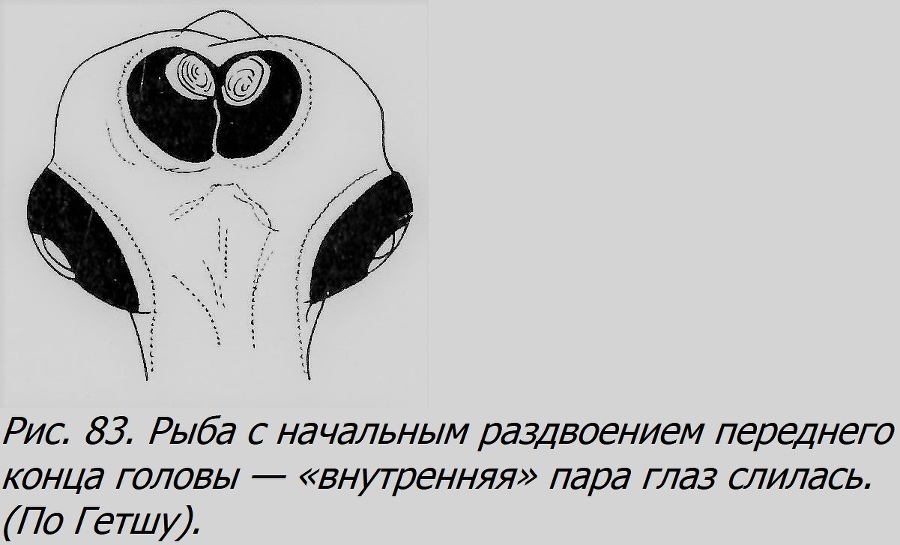
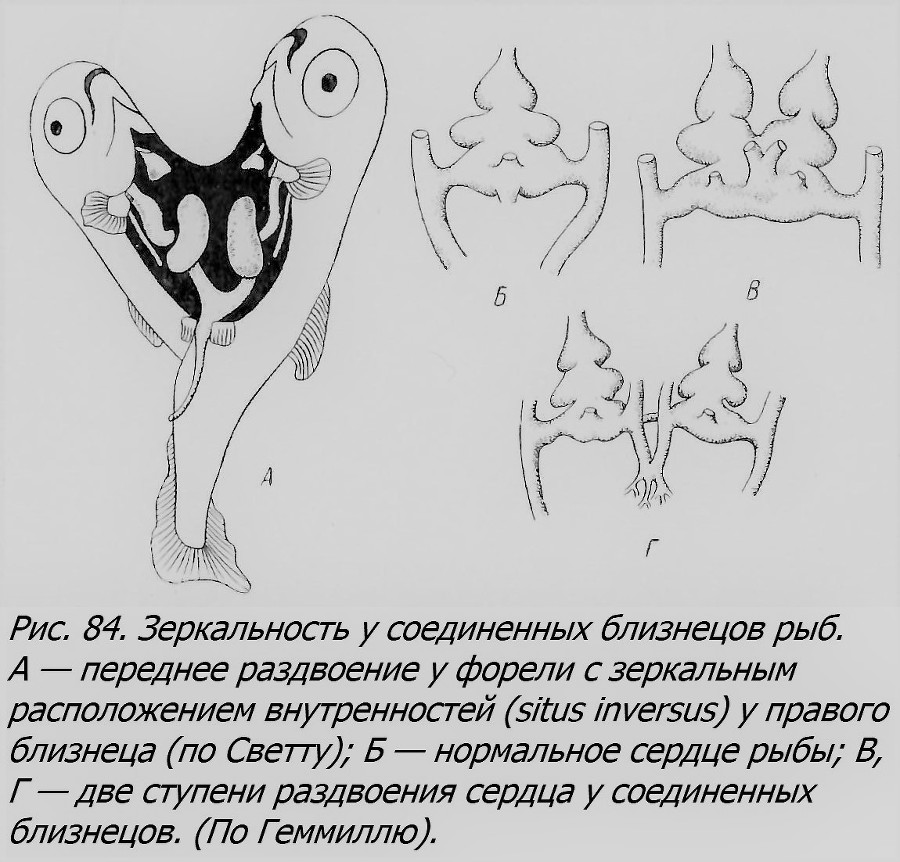
Только что рассмотренный тип соединенных близнецов мы считали, вслед за Стокардом (Stockard, 1921) и другими, результатом вторичного слияния двух первоначально разных зародышей, возникших из одного яйца. Однако такие объекты некоторые авторы, например Ньюмен (Newmen, 1923), принимают за результат продольного расщепления первоначально одного эмбриона, относя их ко второму типу образования близнецов у рыб, о котором речь была выше. По аналогии с другими высшими позвоночными этот тип вероятен и у рыб, тем более, что есть факты, которые, по-видимому, прямо говорят в пользу его существования. Так, например, Бэр (1845а) описал два случая соединенных близнецов окуня, которые он решительно считал за продукты расщепления одного эмбриона, а не слияния двух. Один из этих двойных уродов был менее раздвоен, чем другой (рис. 15). Оба были вполне прозрачны и не имели никаких следов срастания. Бэр сравнивал их с аналогичной стадией раздвоения цыпленка, описанной им (Бэр, 1827). Конечно, в отношении некоторых конкретных объектов может быть окажется не легко решить, каким путем они возникли — первым или вторым. Но во всяком случае, оба варианта являются ОБ.
Близнецы у рыб были получены также экспериментально. Стокард (Stockard, 1921) действовал на ранние стадии развития яиц двух видов рыб — фундулуса и форели — факторами, задерживающими развитие: низкой температурой (до 6—8° С) и уменьшением содержания кислорода в воде, необходимого для интенсивного обмена дробящихся яиц. У обоих видов рыб временная задержка развития вызывала возникновение большего, чем обычно встречается, числа близнецов разного рода. Так, например, у фундулуса при развитии в нормальных условиях одна двойня встречается приблизительно на тысячу яиц, а в эксперименте с пониженной температурой в некоторых случаях число близнецов достигало 9.3% всех яиц. Помимо образования близнецов холод и удушье вызывают гибель значительной части яиц. Также возрастает число различных аномалий развития и уродств. Интересно, что яйца одной кладки одной и той же особи имеют, по-видимому, разную степень сопротивляемости по отношению к вредящим факторам. Одни яйца гибнут, другие дают двойни или уродства, а третьи развиваются нормально. Сущность этих различий еще недостаточно выяснена. Есть также видовые различия реакции на вредные факторы. Форель, например, дает заметно больший процент близнецов, чем фундулус. По-видимому, в яйцах форели легче возникает физиологическая изоляция отдельных участков под влиянием вредящих воздействий, чем в яйцах фундулуса, согласно представлению Стокарда. В зависимости от того, на какой стадии эмбриогенеза действовал вредящий фактор, происходит остановка развития тех органов, которые наиболее интенсивно развиваются на этой стадии, а в связи с этим — соответствующие дефекты развития этих органов (мозга, глаз, конечностей и т. д.). Вредящее, искажающее развитие влияние оказывает также на своего партнера более развитой компонент, автозит на паразита, который в свою очередь тоже не может быть безразличным для автозита.
Другая серия опытов с целью получения близнецов была предпринята на яйцах фундулуса, подвергавшихся минутному облучению ультрафиолетовыми лучами (Hinrichs a. Genther, 1931). Яйца облучались, начиная с 5 минут после оплодотворения. Наибольшее число близнецов получалось под влиянием облучения на 25—30-й минуте и достигало 8.9%. И в этих опытах образование близнецов сопровождалось множественной гибелью яиц и разнообразными уродствами. Облучение за первые полчаса после оплодотворения яиц приходилось на такой период их жизни, когда дробление еще не начиналось или только что начиналось и никакой видимой морфологической дифференцировки еще не было. Большинство близнецов возникало, несомненно,из одного бластодиска, разделившегося, как думают авторы, еще задолго до гаструляции. Образование двух бластодисков на одном яйце с достоверностью не установлено. Раннее облучение способствует раздвоению переднего конца эмбриона, более позднее — заднего, поскольку развитие начинается с головного конца. Интересны некоторые числовые отношения полученных близнецов при облучении не позже, чем через 20 минут после оплодотворения; например, из 106 близнецов было получено: 5 пар с нормальными головами, 21 пара с одной нормальной и одной ненормальной головой, 9 пар, состоящих из слабо дифференцированной массы, одна пара имела частично раздвоенное сердце и 70 пар имели признаки слияния по основной оси (удвоенные ряды сомитов[12] и т. п., — рис. 85). Большинство, таким образом, состояло из соединенных близнецов, получившихся, вероятно, путем слияния двух самостоятельно образовавшихся зачатков, расположенных сравнительно близко друг от друга на общем бластодиске. Гаструляция на противоположных сторонах диска, т. е. под углом 180°, встречается сравнительно редко (рис. 85, Л). Микроскопическое изучение близнецов фундулуса, возникших под влиянием облучения как свободных, так и соединенных, обнаружило зеркальное расположение органов (Hinrichs, 1938). Рисунки, к сожалению, в этой работе не даны, и описание очень кратко; процент s. i. не указан.