Нам остается еще кратко рассмотреть некоторых представителей второго раздела (Б) таблицы Швальбе — несимметричные соединенные образования (duplicitas asymmetros). Основная особенность этой группы заключалется в том, что один из компонентов достигает более или менее полного развития (автозит), тогда как другой в той или иной мере отстает и может жить только за счет своего близнеца (паразит). Один из самых удивительных случаев этого рода был обнаружен в Индии. Это был в общем нормальный ребенок, родившийся 1783 г., на голове которого находилась живая паразитическая голова его недоразвившегося рассосавшегося в материнской утробе близнеца (рис. 45). Рот паразитической головы мог производить сосательные движения и выделять слюну: нос выделял слизь, зрачек сохранившегося глаза (другой пострадал во время родов) мог сокращаться на свету. Во время сна ребенка глаза паразитной головы не были вполне закрыты. Когда ребенок плакал, голова-паразит тоже морщилась и как бы плакала. Когда ребенок сосал, то и паразитическая голова будто бы выражала удовольствие и слюна у нее выделялась обильнее, чем обычно. Когда щипали паразитическую голову, ребенок, видимо, этого не чувствовал или меньше чувствовал, чем когда щипали его собственную голову. Уши, язык и нижняя челюсть паразита были недоразвиты, но зубы появились. Ребенок с паразитической головой был зарисован в возрасте около 2 лет; вскоре он погиб от укуса змеи. Череп его был доставлен в Англию анатому Хантеру (Home, 1790).
В редких случаях автозит, имеющий крупного паразита, достигает зрелого возраста. Примером может служить генуезец Коллоредо (рис. 11), родившийся около 1620 г. и описанный в книге Лицетуса (Licetus, 1665). Паразит висел на туловище автозита головой вниз, имел две недоразвитые руки, каждая с тремя пальцами, и недоразвитую левую ногу. Из открытого рта паразита текла слюна, глаза были закрыты, он иногда слегка шевелил губами.
Другой случай описан в XIX в. Это индус Лалоо, на груди которого находился крупный паразит, лишенный головы, но имеющий четыре конечности. Заднего прохода паразит не имел, но имел пенис, из которого временами выделялась моча. Лалоо прожил свыше 30 лет и был якобы женат (Груздев, 1914, и др.).
Редкие случаи других асимметричных уродов описаны в тератологической литературе (Швальбе и др.) и представляют специальный интерес, почему мы здесь на них останавливаться не будем.
В ряде случаев близнец-паразит деградирует до бесформенной массы тканей, иногда содержащей зуб или участок недоразвитого органа, и образуя род опухоли на теле автозита (рис. 46). Такие опухоли удаляются иногда хирургическим путем, как и более развитые паразиты. Успех такой операции прежде всего зависит от местоположения паразита и степени его слияния с какими-нибудь важными органами автозита.
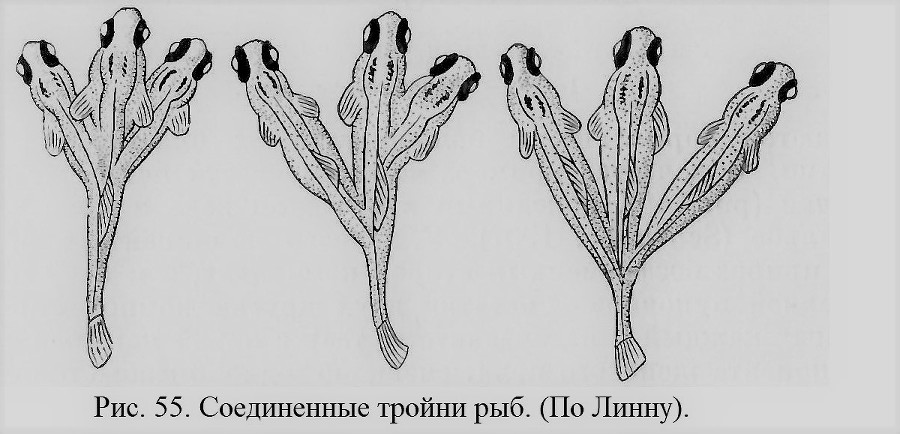
Тройни, соединенные вместе, очень большая редкость (рис. 54). На изображенном уроде видно, что сначала произошло раздвоение переднего конца эмбриона, а затем повторное раздвоение левой (на рисунке) головы. Вскрытие показало, что урод имел 3 гортани, 3 трахеи и 3 пищевода, но 2 пары легких, 2 сердца, 2 позвоночника, из которых один раздвоился вторично только в самом переднем конце. Эта редкая тройня аналогична тройням рыб, изображенным на рис. 55. Некоторые другие тройни изображены в специальных работах (Ahlfeld, 1880; Schwalbe, 1907, и др.).
У животных подобные уродства также представляют большую редкость. Например, описан тройной ягненок, родившийся мертвым. Он состоял из основного, более близкого к норме компонента и двух придаточных, только частично развитых. Голова и шея были нормальные и одиночные. Грудь двойная, 2 пары передних конечностей. Позвоночников было 2 и 2 хвоста, но неравные по размерам. Задних конечностей было 3 пары. Третья пара ног имела рудимент таза и являлась типичным паразитом (Lesbre et Tagant, 1927).
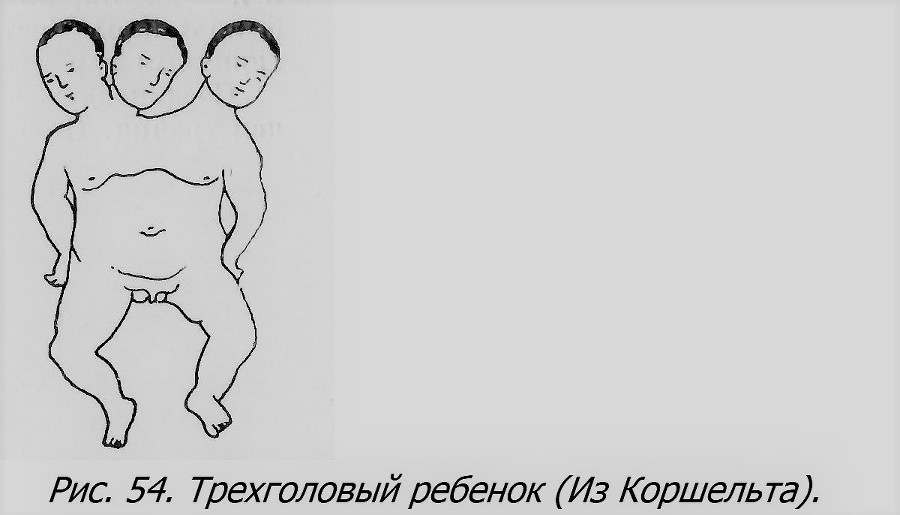
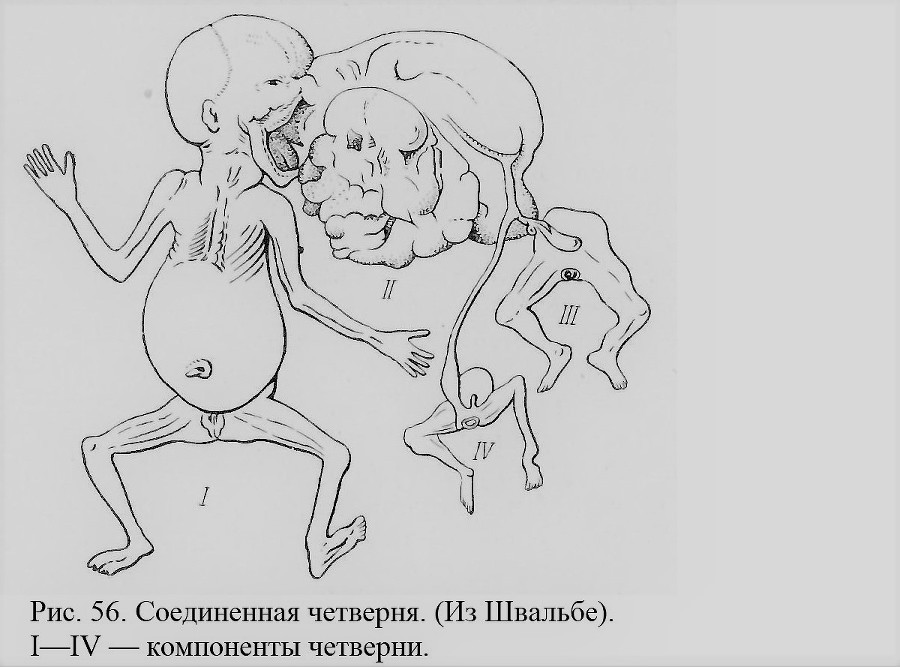
Соединенные четверни, разумеется, встречаются еще реже, чем тройни. По-видимому, компоненты в этих случаях не развиваются нормально, и большинство их оказывается паразитами. В качестве примера можно привести исключительный случай (рис. 56), описанный в прошлом веке и приведенный Швальбе (Schwalbe, 1907). У первого компонента в области рта прирос бесформенный второй компонент, а к нему на раздвоенной пуповине — остатки двух других компонентов, сведенных каждый к недоразвитому тазу с парой ног. Все четыре компонента женского пола, очевидно однояйцевого происхождения. Урод этот погиб в качестве выкидыша на пятом месяце беременности.
Вопрос о способах возникновения соединенных близнецов до сих пор нельзя еще считать вполне выясненным, несмотря на ряд гипотез в этой области, которых выше мы уже не раз касались. Попытка унифицированного объяснения всевозможных форм двойных уродств Грэпером (Graper, 1931) путем сведения их к разнообразным столкновениям «потоков» тканей в период ранней гаструляции, по-видимому, ошибочна. Вероятно, разные типы соединенных близнецов возникают на различных стадиях развития и разными путями (Politzer, 1955).
Прежде всего надо подчеркнуть, что соединенные близнецы являются, очевидно, всегда однояйцевыми. В пользу этого допущения говорит целый ряд фактов. Оба компонента всегда одного пола. У них всегда одинаковы группа крови и факторы крови. Амнион у них один общий. Часто они соединены гомологичными местами груди, черепа и т. д., и компоненты симметрично расположены по отношению друг к другу. Это последнее обстоятельство лежит в основе классификации двойных уродств. Нет ни одного достоверного факта, говорящего в пользу допущения срастания двух эмбрионов, возникших из разных яиц, и образования таким путем двойных уродств (Е. Wolff, 1948, и др.). Экспериментально можно, как известно, достигнуть слияния двух зигот или двух зародышей с образованием гармоничного зародыша удвоенного размера или «сросшихся близнецов», например у морских ежей (Driesch, 1903, и др.) или амфибий (Боги, 1897; Mangold u. Seidel, 1927; Е. Wolff, 1948, и др.). Но это возможно только при удалении оболочек и при других экспериментальных вмешательствах в процессе развития, т. е. таких изменениях его, которые в естественных условиях произойти не могут.
В дальнейшем мы на конкретном материале познакомимся с экспериментальным получением соединенных близнецов у разнятых форм животных, начиная от низших, и объяснениями отдельных случаев такого рода. Здесь же мы лишь кратко остановимся на некоторых общих представлениях об этих явлениях, дополнительно к вышесказанному о причинах возникновения ОБ.
Согласно новейшим представлениям (Politzer, 1955), соединенные близнецы могут возникать по крайней мере тремя путями: 1) во время ранней гаструляции происходит раздвоение переднего конца зародыша; 2) происходит «слияние» тканей в период гаструляции двух смежных зародышей (ОБ) еще до дифференцировки органов; 3) происходит «срастание» двух зародышей (ОБ) уже после известной дифференцировки органов, после гаструляции. Рассмотрим эти три способа образования двойных уродств.
Переднее раздвоение, с которым мы уже не раз встречались, как известно, может быть также получено экспериментально, например перетяжкой на яйцах тритонов. В этих опытах те части тела, которые оказались раздвоенными, получают приблизительно около половины нормально полагающегося материала и оказываются в связи с этим меньшего размера против нормы, а кроме того, иногда еще с дефектами развития (циклопия и т. п.). В случаях образования переднего раздвоения в естественных условиях, т. е. без экспериментального вмешательства, удвоенные передние части тела имеют нормальную величину («закон» Тура, 1903), чем и отличаются от переднего раздвоения, полученного в опыте. Политцер считает, что этим можно объяснить возникновение раздвоения в естественных условиях: вероятно, в силу каких-то физиологических причин (пока не выясненных) подлежащего инвагинации при гаструляции материала оказывается больше, чем полагается, и раздвоение его происходит в порядке регуляции. Хотя закон Тура наблюдается у разных форм животных (амфибий, птиц, человека), однако объяснение его как результат «регуляции» вызывает сомнение и требует более убедительного обоснования, чем у Политцера. Остается неясным также самый «механизм» такого раздвоения при инвагинации. Политцер допускает разные варианты его: или образование двух смежных бластопоров с последующим слиянием тканей внутри гаструлы, как это описано на одном яйце ящерицы (Kopsch, 1897), или раздвоение сначала нормальной инвагинации по пока не выясненной причине. Заднее раздвоение, обычно встречающееся гораздо реже переднего, вряд ли может быть объяснено подобным образом; Политцер предполагает, что этот род раздвоения можно объяснить «срастанием».