Наиболее доступны для кактусовода, далекого от биохимии, первые два, хотя и их раздобыть в чистом виде нелегко.
опытная группа 1 опытная группа 2 контрольная группа
Фото 241 - 243. Опыт по снятию апикального доминирования:
несколько укорененных побегов-деток от одного растения Echinopsis hybridum были объединены в три группы. У кактусов первой опытной группы была удалена верхняя часть стебля, кактусы второй опытной группы были инъецированы 0,0003% раствором зеатина, растения третей группы являлись контролем. На фотографиях показаны растения из 1-й, 2-й и контрольной групп через 6 месяцев после начала опыта. Кактусы из опытной группы 1 (фото 241) претерпели сильнейший стресс, лишившись части стебля, и значительно отстали в нарастании массы стебля от растений 2-й и контрольной групп. Рост основного стебля кактусов второй опытной группы (фото 242) несколько замедлен по сравнению с контролем, однако обильное образование и интенсивный рост боковых побегов в сумме превышает нарастание массы стебля по сравнению с контролем (фото 243).

Фото 244 - 246. Воздействие различных концентраций ауксинов и цитокининов in vitro на клетки каллюса и дифференциация каллюсных клеток при изменении соотношения этих гормонов в питательной среде (по R.Starling):
Astrophytum asterias — рост каллюса и его дифференциация в течение двух месяцев (244);
Leuchtenbergia principis — рост каллюса и его дифференциация в течение двух месяцев (245);
Leuchtenbergiaprincipis — рост каллюса и его дифференциация в течение шести месяцев(246)
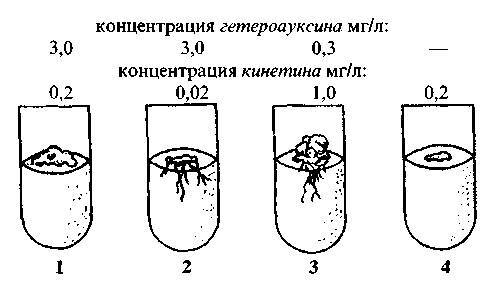
Рис. 68. Повторение опыта Н.И.Якушки-ной по совместному влиянию ауксинов и кинетинов на культуру клеток вторичной меристемы (камбия) взятых от полугодовалых сеянцев Lophophora williamsii.
1 — рост каллюсной массы без заметной дифференциации клеток;
2 — активная дифференциация каллюсных клеток с образованием придаточных корней и началом утолщения корней у основания;
3 — активная дифференциация каллюсных клеток, в основном, с образованием стеблевой части кактуса со многими апексами и лишь незначительными корнями;
4 — очень медленный рост каллюса.
Центром синтеза цитокининов является кончик корня, оттуда эти гормоны разносятся по всему растению. Действие цитокининов обратно действию ауксинов: они снимают эффект апикального доминирования, активизируют рост вторичной меристемы — камбия. Меристема в зоне боковых ареол находится под постоянным воздействием этих двух групп гормонов. Интересно, что на боковую ареолу влияют ауксины, синтезируемые только в апексе стебля и цитокинины, образованные в апексах корней. Чем дальше ареола расположена от верхушки стебля, тем меньшее влияние на нее оказывают ауксины и большее — цитокинины. Как результат — подавляющее большинство деткующихся видов (за исключением кактусов с древовидной и кустовидной формой стебля) начинают образовывать боковые побеги в нижней части стебля. Если же ветвление происходит в верхней части стебля кактуса, то можно с уверенностью сказать, что ауксин-синтезирующая активность апикальной меристемы стебля снижена. Вызвать подобное явление можно и искусственным путем, а не только удалив апекс кактуса. Так апикальное доминирование может быть снято путем обработки точки роста стебля довольно высокими дозами кинетика (1 — 3 мг/л) при инъекции или нанесении ланолиновой пасты.
Цитокинины также задерживают старение растения, являясь, тем самым, антагонистами этилена, ускоряют процессы синтеза белка, стимулируют, как и гиббереллины, прорастание семян и повышают моторику корневой системы.
Однако говорить о полном противопоставлении цитокининов и ауксинов было бы неправильно. В ходе работ отечественных физиологов А.Л.Курсанова, В.И.Кефели, Н.И.Якушки-ной по культивированию меристемы на искусственных средах было установлено, что наибольшего проявления видимого роста тканей, органов, да и всего растения можно добиться лишь при комплексном применении этих гормонов.
Оптимальное соотношение цитокининов и ауксинов для размножения кактусов путем микроклонирования — 0,5 мг/л к 1,8 мг/л соответственно с отклонениями ± 3,1 — 7,3%. При таком сочетании прежде всего стимулируется развитие корневой системы, а затем и всего растения.
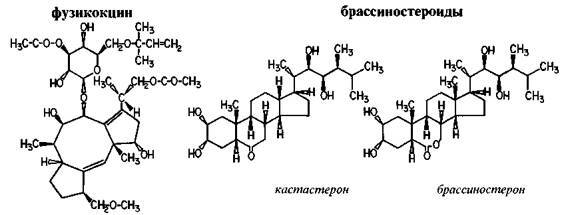
Говоря о гормонах-стимуляторах ростовых процессов, следует хотя бы вскользь упомянуть еще две группы гормональных веществ аналогичного действия: фузикокцин и драссиностероиды. Первый из них — фузикокцин — был выделен итальянскими учеными в начале 60-х годов из патогенного гриба Fusicoccum amygdali, вызывающего гниение персиковых и миндальных деревьев. Этот гормон связывается с мембранами растительных клеток, увеличивает их проницаемость, влияет на синтез белка. Действует фузикокцин аналогично гиббереллинам, но в отличие от последних, стимулирует рост корня и проявляет эффективность при концентрациях 680,8" 10 — 680,8*10"7г/л.
Брассиностероиды продуцирует патогенный гриб Cercospora arachidicola, но впервые они были обнаружены в пыльце рапса (Brassica rapus) — крестоцветного растения, родственного капусте, редису, свекле — за что и получили первую часть своего названия. Вторая часть — «стероиды» говорит о большой близости этих гормонов к стероидным гормонам животных и человека, в частности к половым, поэтому не удивительно, что брассиностероиды обнаруживаются в пыльцевых зернах, хотя в незначительных количествах. Эти гормоны обнаружены в вегетативных
Рис. 69. Фузикокцин и брассиностероиды.
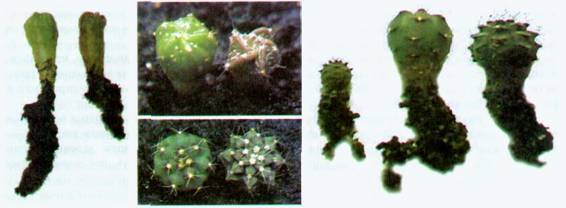
Фото 247 - 250. Воздействие фузикокцина и брассиностероида на сеянцы кактусов: на фотографиях 247 (слева) и 248 (в центре вверху) — сеянцы Astrophytum capricorne в возрасте 5-ти месяцев, левый сеянец обработан фузикокцином. На фотографии 248 (вид сверху) видно изменение габитуса у обработанного сеянца (слева) по сравнению с типовым габитусом одновозрастного растения;
на фотографиях 249 (в центре внизу) и 250 (справа) показаны сеянцы Gymnocalycium pungens. На фотографии 250 (слева направо): трехмесячный сеянец из контрольной группы, трехмесячный сеянец, обработанный брассиностероном и восьмимесячный сеянец, выращенный по методу «тургорной разгонки» без гормональной обработки. Последние два сеянца сфотографированы сверху (фото 249) для показа изменения габитуса обработанного сеянца (слева) по сравнению с типовым габитусом одноразмерного растения.
органах и цветках. Брассиностероиды стимулируют деление и рост клеток, но также и синтез этилена — гормона старения.
При содержании кактусов в культурных условиях кактусоводу, за редким исключением, не приходится задумываться о торможении процессов роста отдельных органов и целого растения. Хотя иногда могут возникать некоторые проблемы, например, как прекратить рост и вытягивание стебля у видов, произрастающих в горах и начинающих расти уже при +10 °С. Поэтому было бы неверно не упомянуть о фитогормонах-ингибиторах физиологических процессов, так как они не только тормозят рост, но и влияют на созревание клеток, подготовку растения к экстремальным условиям (засуха, холод) и т.д.
Этилен — гормон старения. Он участвует в процессах созревания плодов, обуславливает покой почек. Образуясь в околоплоднике, этилен подавляет действие ауксинов, в незначительном количестве синтезируемых в семенах. Таким образом семена, как носители новой жизни, способны долгое время сохраняться при неблагоприятных условиях.
Для растений, как и для их семян, очень важен годовой цикл температуры, влажности, освещенности и т.п. При низких температурах у кактусов снижается интенсивность процессов видимого роста, растения переходят в состояние стагнации. Для семян, находящихся в воздушно-сухом состоянии, низкие температуры играют стимулирующую роль, способствуя ферментативному разложению крахмала и подготовке зародыша к прорастанию. Вот почему кактусам и их семенам необходим период покоя, однако при повышении концентрации этилена во внешней среде длительность этого периода во много раз сокращается (в некоторых опытах практически до нуля).