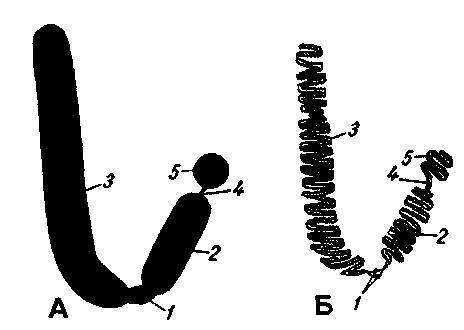
Рис. 117. Строение хромосомы. А. внешний вид Б. внутренняя структура
1. центромера
2. короткое плечо
3. длинное плечо
4. вторичная перетяжка
5. спутник
* этот процесс носит на мание трансляция от латинского translano — передавать.
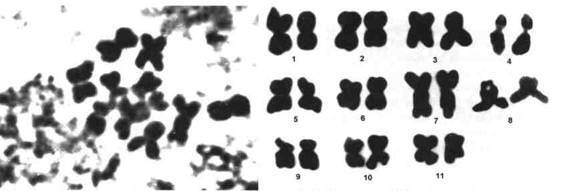
Фото 543. Хромосомы Rhipsalis ramulosa на стадии метафазы.
Фото 544. Кариотип Rhipsalis ramulosa на стадии метафазы. Длина генома 50,9 ± 2,75 мкм, 7-я и 8-я пары хромосом имеют спутники.
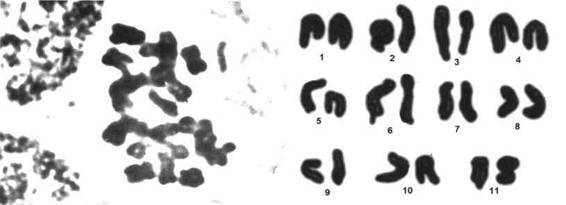
Фото 545. Хромосомы Astrophytum ornatum на стадии метафазы.
(фотографии 543 - 546 В.Панкина)
Фото 546. Кариотип Astrophytum ornatum на стадии анафазы. Длина генома 71,8 + 12,3 мкм, 3-я и 6-я пары хромосом имеют спутники.
После этого рибосома сходит с и-РНК, от нее отделяется полипептидная цепь, и она направляется в начало той же или другой и-РНК.
Как уже говорилось выше, источник наследственной информации — молекулы ДНК, сосредоточены в хромосомах. Хромосомы в период метафазы, о которой будет сказано несколько ниже, представляют собой продолговатые тельца, неравно разделенные перетяжкой — центромерой. Таким образом, в строении хромосом различают центромеру и плечи. Некоторые хромосомы имеют вторичную перетяжку, отделяющую от плеча спутник. Хромосомы имеют отличную друг от друга форму и размер. Набор хромосом, отличающихся друг от друга по внешнему виду, называется гаплоидным (на письме обозначается — п). В ядре вегетативной клетки у каждой хромосомы есть парная хромосома, аналогичная по форме и по размеру, поэтому хромосомный набор вегетативной клетки называется диплоидным (2п), а сам комплекс хромосом, характеризующийся их числом, формой и размерами — кариотипом.
У кактусов гаплоидный набор представлен одиннадцатью хромосомами, диплоидный — 22. Однако существует довольно значительное количество видов, имеющих кариотип, состоящий из гораздо более чем 22 хромосом, соответствующий формуле llm, где т показатель кратности увеличения гаплоидного набора хромосом.
Явление полиплоидии — не редкость в растительном мире, встречается оно и в семействе Cactaceae. Дарлингтон в своих исследованиях определил широкую изменчивость количества хромосом у некоторых кактусов (2п):
Pereskia sacharosa 22
Opuntia salmiana 44
Opuntia ficus-indica 88
Selenicereus pteranthus 22
Astrophytum asterias 22
Echinocactus grusonii 22
Ferocactus acanthoides 22
Hamatocactus setispinus 22
Mammillaria bocasana 22
Mammillaria compressa 44
Mammillaria capensis 264.
Причем, чем более широко по видовому составу представлен род, тем больше случаев полиплоидии можно встретить среди его представителей. Даже в пределах одного ботанического названия или одной популяции встречаются полиплоидные экземпляры. Так, Ремски у некоторых экземпляров Mammillaria prolifera обнаружил кариотипы, доходящие до 2и=66, а у Mammillaria parkinsonii — до 2л=88 хромосом.
На кафедре генетики Московской Сельскохозяйственной Академии им.К.А.Тимирязева В.Х.Панкин провел исследования кариотипов Rhipsalis ramulosa и Astrophytum ornatum. Несмотря на то, что количество хромосом у этих далеких друг от друга в родственном отношении видов одинаково 2л=22, их внешний вид настолько различен, что В.Панкин делает предположение о возможности применения характеристик кариологических данных в определении систематического положения видов.
Для того, чтобы продолжить рассмотрение темы, автору необходимо хотя бы коротко ознакомить читателя с процессами, происходящими при делении клеток. Рост подавляющего большинства живых организмов связан с увеличением численности клеток в тканях путем деления.
Деление клеток происходит в два этапа: деление ядра — митоз* и деление цитоплазмы — цитокинез. Для нас наиболее важно рассмотреть механизм деления ядра и хромосом, в нем расположенных.
Биологический смысл митоза заключается в удваивании числа хромосом и в точном распределении идентичных дочерних хромосом с содержащейся в них генетической информацией между ядрами двух дочерних клеток. Таким образом, суть митоза сводится к двукратному увеличению численности хромосом. Этот способ деления ядра у растений был открыт в 1874 году И.Д.Чистяковым, а детальные исследования были выполнены несколько позже Э.Страсбургером.
В процессе митоза выделяют пять последовательных фаз: профаза, прометафаза, метафаза, анафаза и телофаза.
Непосредственно перед делением ядра белковая оболочка хромосом растворяется и двойная спираль ДНК распадается на две цепочки. Каждая из цепочек ДНК является матрицей для воссоздания парной цепочки. Свободные нуклеотиды занимают сопарные места и соединяются во вторую цепочку. Этот процесс напоминает синтез и-РНК (транскрипцию), с той лишь разницей, что цепочки ДНК расходятся полностью, происходит полное воссоздание сопарной цепочки и, следовательное, удвоение числа первичных молекул ДНК — этот процесс называется репликация. После репликации ДНК каждая хромосома состоит из двух сестринских хроматид. Хроматиды не свертываются в спираль, поэтому в ядре клетки, готовой к делению, видны не отдельные хромосомы, а клубок из тонких слабо спирализированных нитей.
В начале профазы хромосомы начинают скручиваться, вследствие чего укорачиваются и утолщаются. Хроматиды несколько отходят друг от друга, оставаясь связанными лишь центромерами — хромосомы приобретают А'-образную форму. К концу профазы происходит
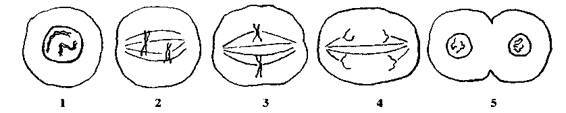
Рис. 118. Схема митоза в гипотетической клетке, содержащей две хромосомы:
1. профаза — произошло удваивание молекул ДНК;
2. прометафаза — оболочка ядра разрушается, и хромосомы прикрепляются к нитям;
3. метафаза — хромосомы сосредоточены в области экватора;
4. анафаза — хромосомы отделяются друг от друга и отходят к полюсам;
5. телофаза — восстанавливается оболочка ядер, и начинается цитокинез.
* митоз — от греческого mltos — пить.
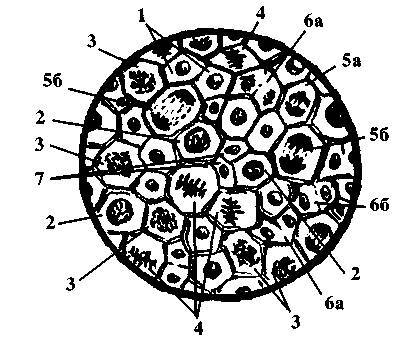
Рис. 119. Клетки апикальной меристемы
сеянца Parodia aureispina:
1. интерфаза;
2. профаза;
3. прометафаза;
4. метафаза;
5а. ранняя анафаза;
56. поздняя анафаза;
6а. ранняя телофаза;
66. поздняя телофаза;
7. молодые дочерние клетки.
растворение оболочки ядра, и хромосомы выходят в цитоплазму. В это же время появляются тонкие нити, отходящие от полюсов клетки к центромерам хромосом — образуется веретенообразная фигура.
В прометафазе цитоплазма в центре клетки разжижается, и хромосомы подходят к области экватора.
В метафазе хромосомы находятся упорядоченно в области экватора. В это время их можно подсчитать, изучить форму.
В анафазе хроматиды полностью отходят друг от друга, превращаясь в отдельно расположенные хромосомы. Нити веретена сокращаются, и хромосомы расходятся к полюсам клетки. Расхождение происходит быстро и одновременно, это хорошо видно под микроскопом в отраженном свете, т.к. в проходящем свете живые хромосомы видны плохо.
В телофазе хромосомы сосредотачиваются у полюсов, деспирализуются и вокруг них восстанавливается оболочка ядра.
Далее происходит деление двуядерной клетки в экваториальной области, образуются две дочерние клетки, вступающие в стадию интерфазы, характеризующуюся количественным ростом.