По тем же законам в меньшей степени представлены, с одной стороны «старые» виды, вытесненные с мест их произрастания более приспособленными формами, и, с другой стороны, «молодые» виды, гораздо лучше «главного» вида приспособленные к конкурентной борьбе, но еще малочисленные. При практически не изменяющихся природных условиях подобная приспособленность выражается, прежде всего, в воспроизводстве: более активном и длительном цветении, большем количестве семян в ягоде, более активной и дружной всхожести семян, большей сохранности семян при лежании. Как положительный фактор в конкурентной борьбе можно считать и небольшие размеры стебля, который при неблагоприятных условиях полностью втягивается под землю и засыпается песком и пылью. Если же учитывать культовую значимость ариокарпусов для местного населения, то следует отметить, что крупные экземпляры гораздо активнее изымались с мест произрастания, т.к. мелкие растения труднее отыскать.
Те, кто сеял семена различных ариокарпусов, кто содержит эти удивительные растения в своих коллекциях, знают, что меньше всего хлопот доставляют A.agavoides и A.kotschoubeyanus — в культуре эти виды охотно цветут в различных регионах земного шара, их семена отличаются наибольшей всхожестью, да и сохранность сеянцев, невосприимчивость ко многим заболеваниям у них выше. Можно предположить, что это «молодые» виды.
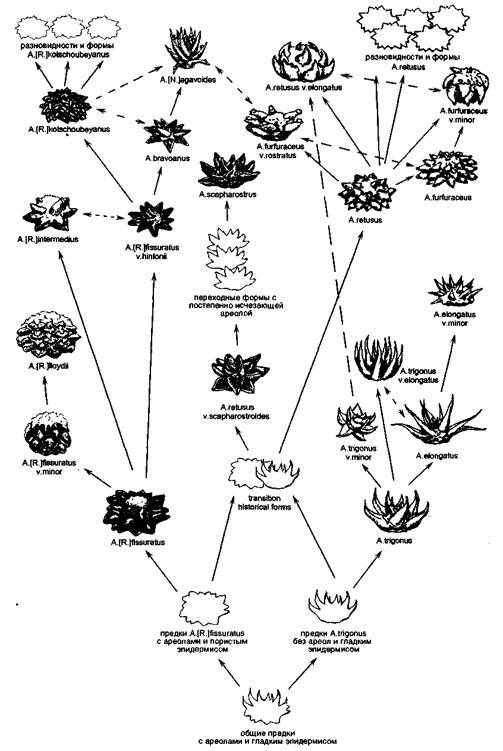
Андерсон, Буксбаум, Крайнц выделяют в роде Ariocarpus 4 группы видов, объединенных по месту расположения точки вегетации и наличию ареолы:
1. точка вегетации расположена в аксилле, ареола отсутствует: A.trigonus, A.scapharostrus;
2. точка вегетации расположена в ареоле: A.agavoides, A.bravoanus;
3. точка вегетации расположена в борозде между аксиллой и ареолой: все представители подрода Roseocactus;
4. точка вегетации расположена в аксилле, но недалеко от кончика сосочка, на его верхней
грани имеется ареола: A. retusus.
Наличие или отсутствие ареолы, хотя бы ее зачатков, как морфологического образования — очень характерный признак. В опытах по скрещиванию видов, имеющих ареолы и лишенных их: A.retusus x A.trigonus, A.retusus x A.scapharostrus, A.agavoides x A.scapharostrus — все гибридные экземпляры первого поколения во взрослом состоянии имели ареолы. То есть признак «наличие ареолы» — доминантный, а «отсутствие ареолы» — рецессивный. Во втором поколении по этому признаку произошло расщепление в соотношении очень близком 3:1.
Другой характерный признак — «пористость эпидермиса» тоже рецессивный, но имеет разную степень выраженности в гетерозиготном состоянии и обуславливается, очевидно, несколькими сцепленными генами. В результате гибридизации A.retusus, имеющего гладкий эпидермис и A.scapharostrus, обладающего пористым эпидермисом, степень пористости эпидермиса в первом поколении была значительно меньше, но не исчезла. При скрещивании экземпляров первого поколения с A. retusus, затем полученных гибридов — опять с A. retusus, степень пористости эпидермиса постепенно снижалась. И только при скрещивании гибридов третьего поколения с чистым A.retusus, т.е. при наличии у гибридных экземпляров 93,5% генов от A.retusus, эпидермис приобрел гладкость. Следует отметить, что во всех гибридных поколениях можно было выделить экземпляры с большей или меньшей степенью выраженности признака.
Вновь обратимся к карте ареала рода Ariocarpus.
Несколько севернее и западнее от мест произрастания A.retusus расположена зона группы A.fissuratus (в которую как близкородственные входят A.intermedius к A.lloydii). Эти растения имеют вегетативную точку, расположенную в бороздке. В верхней части бороздки располагается собственно ареола, из которой у некоторых форм могут появляться мелкие белые колючки. Эпидермис имеет пористый рисунок. Таким образом у A.fissuratus присутствует доминантный признак — «наличие ареолы», и рецессивный — «пористый эпидермис».
Восточнее мест произрастания A. retusus лежит зона произрастания A. trigonus (в которую входитт родственный вид A.elongatus). Эти растения имеют гладкий эпидермис — доминантный признак и отсутствие ареолы — рецессивный признак.
Предположим, пока теоретически, что A.fissuratus nA.trigonus «старые виды». Они наверняка имели общих предков, т.к. показывают большое сходство в строении и форме семян, ягод, цветков, уникальных слизневых каналов и полостей и т.п.
Представим, что предки A.fissuratus и A.trigonus, некогда широко распространенные в современной центральной и северной Мексике «встретились» на территории, которую сейчас занимает A. retusus. В результате многократной гибридизации произошло:
1. выделение доминантного признака «гладкого эпидермиса» в гомозиготное состояние;
2. проявление доминантного признака «наличие ареолы» в полной мере, но удаление вегетативной точки в зону аксиллы;
3. при скрещивании длинно- и короткососочковых родителей, каковыми являлись предки современных A. trigonus и A .fissuratus, у потомков, т.е. у A.retusus, наблюдается средняя длина сосочка.
Рис. 163. Гипотетическое родословное древо рода Ariocarpus (пунктирными стрелками обозначена возможность гибридизации).
Конечно же нельзя говорить, что A.retusus является гибридом A. trigonus w.A.fissuratus, но можно предположить, что предки последних в результате множественных скрещиваний дали начало новым формам, из которых со временем сложилось все современное многообразие разновидностей A. retusus.
Если же сравнить степень воспроизводства гибридов A. retusus xA,trigonus uA.retusus xA.fissuratus с чистыми A returns, взяв показатель воспроизводства последнего за 100%, то средние данные по 10 ягодам от каждого из гибридов составляют 18,81% и 12,02% соответственно. При гибридизации A.trigonus x A.fissuratus средний показатель воспроизводства по сравнению с чистыми A.fissuratus составил 2,77% по 10 ягодам. Полученные гибриды имели удлиненные ареолы и почти гладкий эпидермис, они значительно опережали в росте чистые виды, в привитом состоянии зацвели на третий год и дали множественное расщепление признаков в потомстве.
Среди ариокарпусов можно выделить виды, обладающие высокой степенью пористости эпидермиса. Считается, что этот признак определяется активностью образования кристаллов оксалата кальция — очень гигроскопичных и способных впитывать пары воды из воздуха, даже при ничтожно малой влажности. По другим исследованиям, поры соединяют слизневые полости с атмосферой, в результате чего при неблагоприятных условиях происходит испарение влаги и загустение слизи. При этом стебель значительно сжимается, уходит в грунт. Как правило пористостью эпидермиса обладают виды, наследственно связанные с линией A.fissuratus, имеющей как пористый эпидермис, так и ареолу или бороздку.
Однако у A.scapharostrm, несмотря на сильную степень пористости эпидермиса, наблюдается большее сходство с A.trigonus как по признаку отсутствия ареолы, так и по аналогичному расположению вегетативной точки в субаксиллярном пространстве. В коллекциях и, по некоторым данным, в природе существуют растения, напоминающие A.scapharostrus чрезвычайно пористым эпидермисом и характерной формой сосочков, но имеющие небольшую удлиненную ареолу. Подобные растения иногда именуют A.retusus v.scapharostroides. Если этот кактус действительно существует в природе, то он является переходной формой к A.scapharostrus, в противном случае, пористый эпидермис последнего развился конвергентно как ответная реакция на условия внешней среды.
Ariocarpusflssuratus имеет на верхней грани сосочка характерную U- или F-образную борозду. У A.agavoides подобной борозды нет, но у некоторых экземпляров имеются хорошо заметные выросты по бокам от ареолы. Сравнительно недавно открытые A.fissuratus v.hintonii и A.bravoanus представляют собой переходные формы по признакам «место расположения ареолы» и «наличие V-образной борозды на верхней грани сосочка» от A.fissuratus к A.agavoides. Следует отметить, что все четыре наименования имеют четко выраженный пористый рисунок эпидермиса.