Если же в гетерозиготе встречаются гены фиолетовой и розовой окраски, то цветок будет фиолетового цвета.
При внимательном рассмотрении гибридных цветков можно обнаружить разницу в форме лепестков, интенсивности окраски частей лепестка: краев или центральной жилки и т.п. То есть в данном случае множественные аллели корреллируют с несолькими признаками — являются плейотропными.
Нередки случаи, когда у различных видов, разновидностей или форм один и тот же признак является результатом проявления различных генов. Например, макушечный и боковой цефалий различных родов образованы конвергентно. Даже очень похожие по месту расположения цефалий кактусов из родов Melocactus и Discocactus обусловлены различным сочетанием генов.
Другой пример, признак «нудальности стебля» у близкородственных кактусов Astrophytum myriostigma f.nudum и A.myriostigma v.strongylogonum subv.nudum хотя и является в обоих случаях рецессивным, но определяется сочетанием различных по количественному и качественному составу групп генов. Так, при аналитическом моногибридном скрещивании инцухт-линейных A.mvnVttrigmc/ у. A.myriostigma v.strongylogonum subv.nudum во втором поколении наблюдалось расщепление 3 : 1, в то время как у гибридов A.myriostigma x A.myriostigma f.nudum во втором поколении расщепление признака было примерно 18:1, причем все нудальные формы имели мелкие крапины на грани ребра. Более того, при самоопылении экземпляров Astrophytum myriostigma f.nudum, совершенно лишенных крапа, в потомстве встречаются экземпляры с крапинами. В практике автора подобное расщепление признака проявлялось даже в пятом поколении. При самоопылении A.myriostigma v.strongylogonum subv.nudum в первом и последующих поколениях автору не встречались экземпляры с крапом, что дало возможность вывести инцухт-линию по признаку нудальности стебля.
Предварительно рассмотрев строение молекулы ДНК, принцип линейного расположения наследственных факторов, необходимо уточнить, что весь количественный состав генов делится на определенные группы. Число генов в каждой группе определяет длину первичной структуры молекул ДНК и, следовательно, размер хромосом. Гены, расположенные в одной хромосоме, называются группой сцепления. Поэтому при рассмотрении законов Менделя правильнее было бы оговориться, что эти законы приемлемы, когда гены, определяющие изучаемые признаки, находятся в разных хромосомах, в разных молекулах ДНК, относятся к разным группам сцепления.
Однако, при клеточном делении с образованием гаплоидных половых клеток — мейозе, на определенных стадиях, когда гомологичные хромосомы переплетаются, происходит обмен их участков. Такой процесс носит название «кроссинговер» или .«перекрещивание» хромосом, причем обмен может захватывать как несколько нуклеотидов, так и несколько генов, расположенных в сопарных молекулах ДНК. Кроссинговер может произойти как на одном, так и на многих участках
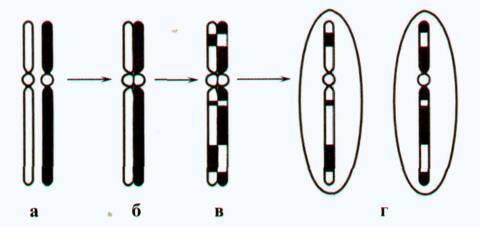
Рис. 155. Принцип кроссинговера:
а — гомологичные хромосомы; б — в профазе I мейоза хромосомы соединены по всей длине; в—кроссинговер—обмен участками хромосом;
г — расхождение хромосом в телофазе и образование дочерних клеток
Фото 974. Пример экспрессивности признака «крапчатости» у A atrophy turn my riostigma: при недостаточном солнечном освещении крап, выполняющий функцию защиты стебля от чрезмерной инсоляции, не образуется, но наблюдается интенсивный рост стебля. При повышении уровня естественного освещения скорость роста стебля снизилась, но возобновилось образование крапа.
Фото 975. Вариабельность проявления признака «крапчатость стебля» у трехгодовалых сеянцев Astrophytum asterias.
Фото 976. Вариабельность проявления признака «количество ребер» у одновозрастных сеянцев Astrophytum cv. Onzuka. (фото В.Гапона)
одной хромосомы, причем чем дальше в одной хромосоме расположены участки, тем чаще можно ожидать между ними перекрещивание. Таким образом, несмотря на то что половые клетки имеют гаплоидный набор хромосом, хромосомы будут нести наследственную информацию как от первоначальных отцовских, так и от первоначальных материнских гамет. Все это ведет, с одной стороны, к большой перекомбинации наследственного материала, но, с другой, сохраняет в потомке информацию о всех его предках.
Как уже было отмечено, согласно современным исследованиям гены делятся на структурные, несущие информацию о строении белков, и функциональные, осуществляющие регуляцию различных биологических реакций. Таким образом, проявление какого-либо признака зависит не только от основных веществ, участвующих в реакциях, но и от скорости и интенсивности этих реакций, от наличия или отсутствия промежуточных продуктов реакций и т.д. Понятно, что здесь не последнюю роль играют условия окружающей среды. Так, замечено, что степень интенсивности окраски цветков зависит от содержания азота в почве, величина колючек — от влажности субстрата и воздуха, толщина эпидермиса — от температуры. То есть один и тот же ген или группа генов могут определять признак довольно вариабельный, изменяющийся в зависимости от параметров внешней среды. Степень фенотипического проявления гена в зависимости от условий внешней среды называется — экспрессивность.
Экспрессивность проявления признаков можно наблюдать и у кактусов. Причем при более мягких условиях культивирования снижается экспрессия ксерофитного фенотипа, характеризующегося специфическим опушением, колючками, наличием развитой кутикулы, размером стебля, но повышается экспрессия мезофитного фе«отипа, прежде всего выраженная в увеличении объема клеток, в сочно-зеленом эпидермисе, снижении степени опушенности и т.д.
Характерный пример различной степени проявления генов могут представлять типичные экземпляры Astrophytum myriostigma, содержащиеся в условиях недостаточного солнечного освещения. При этом в апикальной зоне почти полностью пропадает опушение-крап. Когда же интенсивность естественного освещения увеличивается — крап в апикальной зоне образуется вновь.
Если учитывать, что компоненты внешней среды оказывают влияние на проявление признака не непосредственно, а через изменение интенсивности ряда биохимических реакций, в свою очередь обусловленных наличием или отсутствием специфических белков, то следует сделать вывод, что экспрессивность признака имеет разную степень выраженности, зависящую от конкретного генотипа, возраста растительного организма, его физиологического состояния. Например, в посеве семян даже из одной ягоды у сеянцев можно наблюдать разницу в росте, интенсивности опущения ареол или крапа, количестве ребер, цвете и числе колючек и т.д. С возрастом высока вероятность выравнивания степени проявления признаков: образуются характерные по цвету и количеству колючки, появляется стандартное число ребер, опушение ареол или крапа приобретает типичную интенсивность.
Степень проявления признака в популяции выражается в количественном отношении особей, имеющих этот признак, к общему числу экземпляров в данной популяции. Эта величина называется пенетрантность. Термины экспрессивность и пенетрантность были введены в 1927 г. русским ученым Н.В.Тимофеевым-Ресовским.
Пенетрантность и экспрессивность признака следует учитывать как при селекционной работе, так и при попытке присваивания новых таксономических названий. Очень часто в каталогах фирм, реализующих кактусы и их семена, можно встретить различные наименования, по сути, обозначающие одни и те же растения, но с разной степенью проявления какого-либо признака. При выращивании же подобных «новых» кактусов в культурных условиях различия часто исчезают, и признаки могут проявляться в равной степени.