По химическому строению хлорофилл весьма близок к красному пигменту крови — гемму, но содержит не атом железа, а атом магния. Хлорофилл а отличается от хлорофилла b только тем, что у второго метильная группа заменена на радикал -СОН.
Для образования хлорофилла в растениях необходимо несколько четко определенных условий: присутствие пропластид, способных к позеленению (будущих хлоропластов), света и солей железа, являющихся катализаторами энергопоглотительных процессов, при отсутствии которых наблюдается побледнение (хлороз) зеленых тканей растения.
В зеленых клетках листьев мезофитных растений в результате фотосинтетической реакции образуются простые сахара (глюкоза, фруктоза), которые затем синтезируются в крахмал. Химизм фотосинтеза довольно сложен и представляет собой ряд превращений соединений углерода и образование богатых энергией органических соединений фосфора, т.е. происходит накопление энергии в виде химических связей.
Все реакции фотосинтеза условно делятся на две группы: световые и темповые.
Световые реакции происходят под действием энергии солнца. К ним относятся: разложение воды на кислород и водород, образование энергетических фосфорных соединений и синтез аминокислот и белков.
Темповые реакции проходят (не обязательно ночью и в темноте) под действием энергии химических связей и имеют конечным результатом образование простых Сахаров и выделение в атмосферу молекулярного кислорода.
Оптимальная температура процессов фотосинтеза лежит в пределах +20 — +30 °С. У мезофитных растений в вегетационный период днем идет поглощение углекислого газа, синтез органических веществ и выделение кислорода. Процесс дыхания, который по своей сути обратеи фотосинтезу, проходит постоянно в течение суток, но его интенсивность изменяется в связи с суточными колебаниями температуры окружающей среды. Температурный оптимум дыхания значительно выше, чем фотосинтетический, поэтому на свету при температуре +15 +30 С синтез преобладает над распадом. Если же температура воздуха увеличивается до +45 +50 °С, фотосинтез почти прекращается, в то время как процесс дыхания идет полным ходом. То есть одной из причин угнетения и гибели мезофитных растений при повышенной температуре является их истощение при активном дыхании и угнетении фотосинтеза.
В ходе изучения процессов фотосинтеза было установлено, что для восстановления одной молекулы СО, требуется 12 квантов света, причем только 4 сохраняются в виде химической энергии, остальные 8 используются для образования неустойчивых высокоэнергетических продукт».
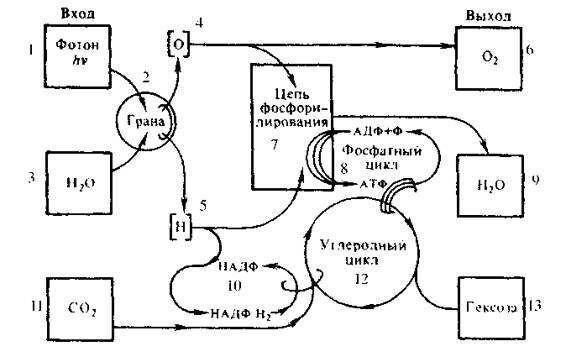
Рис. 37. Блок-схема фотосинтеза:
Под действием кванта света (1) в гранах (2) происходит разложение молекулы воды (3) на кислород (4) и водород (5). Кислород частично выделяется в атмосферу в чистом виде (6), а частично вместе с водородом (5) включается в цепь фосфорилирования (7). Аденозиндифосфат (АДФ) присоединяет атом фосфора (Ф). переходит в АТФ и включается в фосфатный цикл (8). В качестве побочного продукта реакций фосфорилирования выделяется вода (9). Остаток атомов водорода (5) участвует в восстановлении НАДФ в НАДФ«Н (10). НАДФ»Н. АТФ и поглощенный из ат мосфсры углекислый газ (1 1) участвуют в синтезе сахаров (углеродный цикл Калышна) (12). В качестве продукта цикла Кальвина синтезируются шестиуглсродные сахара (гексозы) (1 3). которые необходимы для поддержания энергетики фотосинтеза. В конечном счете эта энергия переходит в тепло. Теперь весьма упрощенно разберем механику поглощения солнечной энергии растениями.
Для поддержания энергетического хода фотосинтеза требуются уже упомянутые высокоэнергетические органические соединения: аденозинтрифосфат (АТФ, аденил-пирофосфорная кислота) и восстановленный водородом никотинамидадениндинуклеотидфосфат (НАДФ). Естественно, довольно трудно выговорить эти длинные названия, поэтому пользуются аббревиатурой.
АТФ и НАДФ*Н образуются во время световой реакции фотосинтеза, т.е. они как бы аккумулируют большую часть солнечной энергии. Процесс усвоения солнечной энергии носит название фотофосфорелирование. Электроны в хлоропластах передаются от возбужденного хлорофилла через ряд органических веществ к месту фотофосфорелирования.
При темновой реакции фотосинтеза происходит цикличное ферментное превращение углерода из двуокиси в сахара при непосредственном участии АТФ и НАДФ*Н. Этот цикл был открыт М.Кальвином и назван в его честь.
Молекула углекислого газа присоединяется к пятиуглеродному фосфорсодержащему сахару и образует шестиуглеродное соединение, которое затем разлагается до трехуглеродных кислот и сахаров, которые, в свою очередь, синтезируются в шестиуглеродные сахара (гексозы) и затем - в крахмал. Смысл такой «кольцевой» реакции сводится к поэтапному переходу энергии из легко разрушаемых высокоэнергетических соединений в более стабильные в энергетическом отношении сахара и крахмал. Такой вид метаболизма называют С,-метаболизм углерода, а мезофитные растения — С,-растениями. Усвоенный из воздуха углерод в дальнейшем участвует в синтезе аминокислот, жиров, углеводов. Обратимся теперь к кактусам.
В аридных условиях в дневное время растения испытывают дефицит в воде. Ввиду того, что снижается обводненность эпидермиса, закрываются устьичные щели, т.е. углекислый газ не может проникнуть в хлоропласты. В ночное время устьица открываются, углекислый газ попадает в клетку и поглощается фосфорсодержащей органической кислотой. В это же время крахмал разлагается до шестиуглеродпых Сахаров, которые, в свою очередь, разлагаются до четырехуглеродной яблочной и изолимонной кислот. Днем накопленный в тканях за ночь углекислый газ включается в состав органических веществ, четырехуглеродные соединения синтезируются в шестиуглеродные, а затем — в крахмал. Параллельно в дневное время происходит активное накопление солнечной энергии в фосфорорганических соединениях АТФ и НАДФ»Н. Подобный тип метаболизма принято называть С4-метаболизмом.
Рис. 38. Схема фотосинтеза у растений с С,- и С4-типом метаболизма (из S.Brehme).
Такой тип превращения органических веществ присущ, кроме суккулентных растений, некоторым злаковым (кукуруза, сахарный тростник, просо, сорго и др.) с той лишь разницей, что процесс фотосинтеза протекает в клетках паренхимы сосудистых пучков, т.е. в более утолщенных частях листовой пластинки. У кактусов и других суккулентов фотосинтетические реакции проходят в зеленом эпидермисе, а избыточное количество крахмала, который, кстати, отрицательно влияет на интенсивность фотосинтеза, выделяется в межклеточное пространство и транспортируется до сосудистых пучков. Этот способ метаболизма, особенно в зарубежной литературе, выделяют из общего С4-метаболизма и именуютт крассуловым типом метаболизма: САМ — Crassuluccan acid metabolism (по латинскому названию растений из семейства Толстянковых, которые исследовали австралийские ученые М.Хетч и С.Слэк).
С4-растения характеризуются мелкими клетками, густой цитоплазмой, повышеннымсинтезом и накоплением органических веществ, менее интенсивным световым дыханием, отсутствием обратного потока углекислого газа в атмосферу при дыхании, т.к. последний абсорбируется в паренхиме запасающей ткани и вступает в дальнейшие реакции фотосинтеза. Таким образом, коллекция кактусов, содержащихся в комнатных условиях, поглотит из атмосферы углекислый газ и обогатит ее кислородом (следует учитывать, что лиственные растения в ночное время выделяют двуокись углерода).
Свет с одной стороны — это поток энергетических частиц — квантов, а с другой — электромагнитные волны. Видимый солнечный свет по качественному составу не однороден. Он образован сложением волн семи цветов (в доказательство этому достаточно вспомнить радугу): красный, оранжевый, желтый, зеленый, голубой, синий и фиолетовый, которые образуют спектр белого цвета. Красные лучи имеют большую длину волны. От красного к фиолетовому цвету