Значительно эволюционировали и общие представления об отношении генов к определяемым ими признакам. Мендель и первые менделисты склонялись к полному отождествлению гена с признаком и надеялись «разложить» каждый организм на сумму совершенно независимых признаков, число которых равно количеству наличных генов. В дальнейшем было установлено, что один ген может определять совокупность признаков, и, наоборот, каждый признак организма зависит от ряда генов (см. Плейотропия ). Т. о., раздельны и независимы при наследовании только гены, признаки же организма следует рассматривать не как мозаику отдельностей, а как единое целое, возникающее в результате развития в конкретных условиях среды. Изучение сложных закономерностей развития наследственно обусловленных признаков составляет предмет самостоятельной науки — феногенетики . Успехи генетики и особенно раскрытие механизмов наследственности на молекулярном уровне (см. Молекулярная генетика ) окончательно упрочили М. как учение об основных закономерностях наследственности.
Менделизм и дарвинизм. М. и сложившаяся в начале 20 в. мутационная теория вызвали сначала антагонизм между «ортодоксальными» дарвинистами и менделистами. Так, Х. Де Фриз полагал (1901—03), что одной «прогрессивной» мутации достаточно для возникновения нового вида. Я. Лотса (1912—13) выдвинул необоснованную теорию неизменности генов и постоянства их числа. Бэтсон (1914) на основании теории «присутствия — отсутствия» утверждал, что наряду с перекомбинированием генов идёт процесс их потери, выпадения. Все эти теории не учитывали, что одни закономерности наследственности не могут объяснить эволюционного процесса. В то же время и многие дарвинисты ошибочно полагали, что эволюция может идти только на основе непрерывных, незначительных изменений, имеющих массовый характер. Они враждебно встретили М. и мутационную теорию, пытаясь отрицать всеобщность установленных этими учениями закономерностей. В резкой оппозиции к М. оказались английские дарвинисты (А. Уоллес , Э. Рей Ланкестер и др.). Основываясь на положениях т. н. биометрической школы (см. Биометрия ), они отрицали прерывистый характер наследственной изменчивости, альтернативную, дискретную наследственность, а также возможность сохранения при свободном скрещивании единичных мутаций. Взгляды английских ортодоксальных дарвинистов повлияли и на некоторых русских учёных, в том числе на К. А. Тимирязева и М. А. Мензбира . Тем не менее Тимирязев понимал, что М. «... служит только поддержкой дарвинизму, устраняя одно из самых важных возражений, когда-либо выдвинутых против него» (Соч., т. 7, 1939, с. 236). Позднее этого не учли и противники М. в СССР.
Согласно господствовавшему до М. учению о «слитной» наследственности, признаки скрещивающихся организмов «сливаются», так что всякий новый, единичный признак не имеет шансов сохраниться в массе отличающихся от него форм вида. Поэтому считалось, что естественный отбор бессилен сохранить подобный признак, даже если он окажется полезным в борьбе за существование . М. позволил отклонить учение о слитной наследственности и вместе с ним эти возражения против теории естественного отбора. Всякий вновь возникший наследственный признак при скрещивании может в ближайшем поколении не проявиться; это, однако, не означает, что определяющий его наследственный фактор «слился» или навсегда «поглотился» в популяции . Признаки, определяемые рецессивными генами, находящимися в гетерозиготном состоянии, могут вновь проявиться при переходе в гомозиготное состояние спустя любое число поколений. Эти представления, естественно вытекающие из М., были теоретически и экспериментально обоснованы С. С. Четвериковым (1926) и его сотрудниками. Независимо от них закономерности эволюционной генетики разрабатывали английские учёные Дж. Б. С. Холдейн (1924 и позже) и Р. А. Фишер (1928—30) и американский учёный С. Райт (1931). К 30-м гг. 20 в. генетика и лежащий в её основе М. стали признанным фундаментом современного дарвинизма.
Т. о., М. сыграл революционизирующую роль в биологии, доказав, что наследственные факторы имеют корпускулярную, дискретную природу, а их переход от поколения к поколению определяется вариационно-статистическими закономерностями. Эти новые принципы позволили разрешить трудности, стоявшие перед дарвинизмом, и разработать современное учение о процессах микроэволюции . М. стал теоретической основой современных методов селекции микроорганизмов, культурных растений и домашних животных, а также вызвал развитие генетики медицинской . См. также статьи Генетика , Дарвинизм , Наследственность и литературу при них.
Лит.: История менделизма — Гайсинович А. Е., Зарождение генетики, М., 1967; Мендель Г., Нодэн Ш., Сажрэ О., Избранные работы, [2 изд., М., 1968]; Zirkle С., The beginnings of plant hybridization, Phil., 1935; Stubbe Н., Kurze Geschichte der Genetik bis zur Wiederentdeckung der Vererbungsregein Gregor Mendels, 2 Aufl., Jena, 1965; Roberts Н. F., Plant hybridization before Mendel, N. Y. — L., 1965; Olby R. C., Origins of Mendelism, L., 1966.
Изложение менделизма — Корренс К., Новые законы наследственности, пер. с нем., М., 1913; Пеннет Р. К., Менделизм, М. — Л., 1930; Богданов Е. А., Менделизм или теория скрещивания, М., 1914; The mechanism of mendelian heredity, rev ed., N. Y., 1926; Bateson W., Mendel's principles of heredity, 4 ed., Camb., 1930; Plate L., Vererbungsiehre, 2 Aufl., Bd 1, Jena, 1932; Ford Е. В., Mendelism and evolution, 7 ed., 1960.
А. Е. Гайсинович.
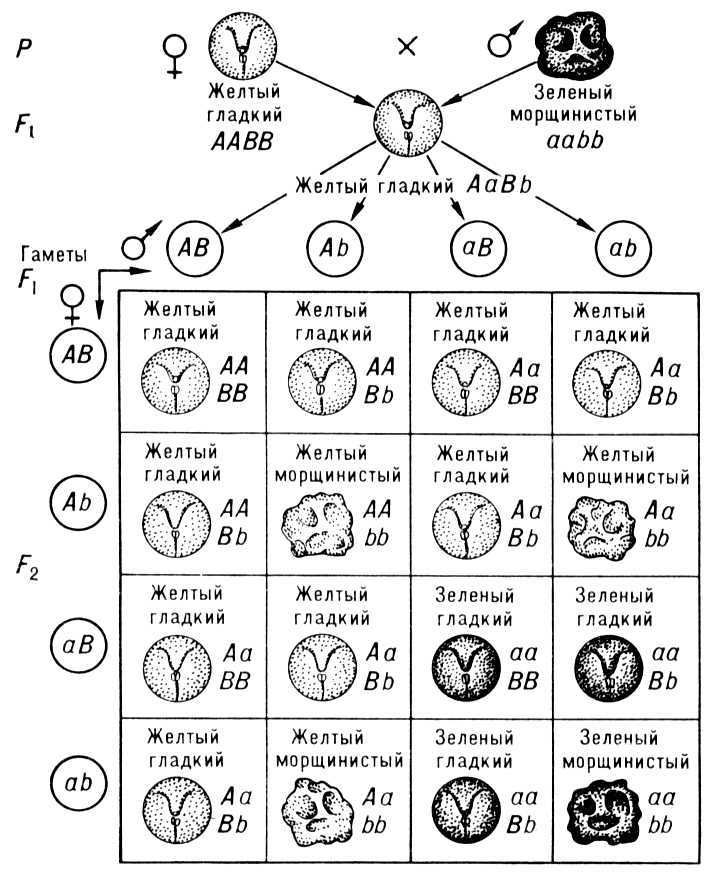
Свободное комбинирование двух пар признаков (окраска и форма семян у гороха) с расщеплением 9 : 3 : 3 : 1; А — жёлтая окраска семян (доминантная), а — зелёная окраска семян (рецессивная); В — гладкая форма семян (доминантная); b — морщинистая форма семян (рецессивная); Р — родительские формы; F — гибриды первого поколения; F2 — гибриды второго поколения.
Мендель Грегор Иоганн
Ме'ндель (Mendel) Грегор Иоганн (22.7.1822, Хейнцендорф, Австро-Венгрия, ныне Гинчице, Чехословакия, — 6.1.1884, Брюнн, Австро-Венгрия, ныне Брно, Чехословакия), основоположник учения о наследственности, названного в его честь менделизмом . Сын крестьянина; в 1843 по окончании философских классов при университете в Ольмюце в связи с материальными трудностями постригся в монахи Августинского монастыря в Брюнне (с 1868 настоятель этого монастыря). С 1849 преподавал в средней школе естественную историю и физику. В 1851—53 вольнослушатель Венского университета, где изучал физику, ботанику, палеонтологию и аналитическую химию. В 1856—63 М. провёл обширные опыты по гибридизации 22 сортов гороха. Результаты этих опытов были доложены им в 1865 в Брюннском обществе естествоиспытателей и опубликованы в «Записках» того же общества (1866). Количественный учёт всех типов полученных гибридов, а также вариационно-статистический подход, характерный для всего склада мышления М., позволили ему впервые обосновать и сформулировать закономерности свободного расхождения и комбинирования наследственных факторов. Эти закономерности легли в основу учения о наследственности и получили название Менделя законов . М. пытался подтвердить обнаруженные им закономерности на др. растениях, в том числе на ястребинке. Выбор объекта оказался неудачным: полученные результаты противоречили установленным на горохе закономерностям. (Как выяснилось позднее, ястребинка часто размножается без оплодотворения, и попытки получения у неё гибридов остаются безуспешными.) М. занимался также пчеловодством, метеорологией, садоводством (вывел новый сорт фуксии, осуществлял прививки и скрещивания плодовых деревьев), скрещивал серых и белых мышей.