Стеариновая кислота
Стеариновая кислота, октадекановая кислота, CH3 (CH2 )16 COOH, одноосновная насыщенная карбоновая кислота алифатического ряда. Бесцветные кристаллы, tnn 69,6 °С, tkип 376,1 °С; нерастворима в воде, растворима в эфире. С. к. является одной из наиболее распространённых в природе высших жирных кислот; глицериды С. к. — главная составная часть многих жиров и масел, из которых её выделяют гидролизом (обычно в виде стеарина — смеси С. к. и пальмитиновой кислоты ). С. к. можно получить дробным осаждением или дистилляцией из стеарина, гидрированием олеиновой кислоты и др. способами. Щелочные соли С. к. являются мылами . Применяют С. к.: очищенную — в органическом синтезе, аналитической химии (для определения Ca, Mg, Li), техническую — как диспергатор ингредиентов и активатор вулканизации в производстве резины. Стеараты натрия, лития, кальция, свинца и др. металлов используют как компоненты пластичных смазок , С. к. и её эфиры — при получении косметических средств.
Стеатитовая керамика
Стеати'товая кера'мика, изделия и материалы, применяемые как изоляторы в высоковольтной и высокочастотной технике; изготовляется на основе минерала стеатита (разновидность талька). С. к. характеризуется значительной прочностью при статическом изгибе — до 190 Мн/м2 (1900 кгс/см2 ), диэлектрическая проницаемость колеблется в пределах 5,5—7, диэлектрические потери (при частоте 1 Мгц и температуре 20°С)(3— 25) 10-4 Изделия из С. к. формуют методами керамической технологии (прессование, литьё под давлением и др.) и обжигают при температуре 1200—1300° С. производство изделий из С. к., особенно крупных размеров, сопряжено с трудностями, обусловленными узким интервалом спекания (10—40 °С). Недостаток С. к. — склонность к «старению» при длительной эксплуатации.
Стеатопигия
Стеатопиги'я (от греч. stear, родительный падеж steatos — жир и pyge — огузок, ягодицы), сильное развитие подкожного жирового слоя у человека на ягодицах (в области большой ягодичной мышцы). Наиболее выражена С. у женщин некоторых южноафриканских народов, главным образом у бушменов и готтентотов . У этих народов, а также у зулу С. считается признаком женской красоты. Причина возникновения С. окончательно не выяснена.
Стебель
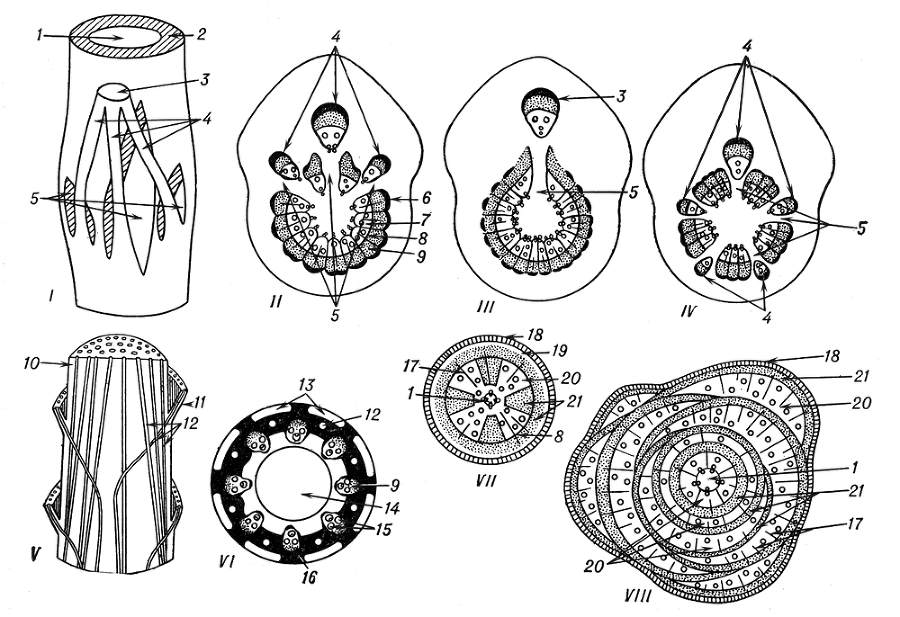
Сте'бель (caulis), осевой орган высших растений, вместе с листьями составляющий побег , служит для передвижения воды и веществ между корнями и листьями, для увеличения ассимилирующей поверхности растения путём ветвления и упорядоченного расположения листьев, а также цветков и плодов; может участвовать в накоплении воды и запасных питательных веществ, в фотосинтезе . Участки С., от которых отходят боковые органы (ветви, листья и др.), называются узлами, участки между узлами — междоузлиями. С. бывают травянистыми и деревянистыми; главный С. древесных растений называется стволом . Форма С. разнообразна: цилиндрическая (наиболее распространена), трёхгранная (осоки), четырёхгранная (губоцветные), многогранная, уплощённая (кактусы) и др. По положению в пространстве различают С. прямостоячие, лежачие, ползучие, лазающие и др.; надземные и подземные (см. рис. 2). Длина С. от 1—1,5 мм (пресноводная вольфия) до 200—300 м (тропические пальмы-ротанги), диаметр от долей мм (мхи) до 10—11 м (баобаб, секвойя). С. растет в длину за счёт деятельности верхушечной меристемы побега, составляющей конус нарастания . Кроме верхушечного роста, у некоторых растений в основании междоузлий происходит ещё интеркалярный (вставочный) рост (например, у злаков).
В С. выделяют анатомо-топографические зоны: наружную — эпидермис, внутреннюю — центральный цилиндр, или стелу, и расположенную между ними зону первичной коры , внутренний паренхимный слой которой превращен в эндодерму. Последняя граничит с периферической зоной стелы (представленной паренхимной или механической тканями) — перициклом (у некоторых растений его нет). Большая часть стелы состоит из проводящих тканей , флоэма находится снаружи от ксилемы . У лиственных мхов в центре С. расположен «проводящий пучок», элементы которого лишь внешне сходны с проводящими элементами флоэмы и ксилемы. У сосудистых растений формированию проводящих тканей предшествует развитие прокамбия . У плаунов ксилема разделена на лентовидные тяжи, окруженные флоэмой, сердцевины нет. У хвощей закрытые коллатеральные пучки с т. н. каринальной полостью вместо ксилемы располагаются вокруг центральной воздушной полости. У папоротников проводящие ткани кольцом окружают сердцевину. В С. семенных растений встречаются пучковый и сплошной типы строения проводящей системы, пересечённой радиально расходящимися паренхимными сердцевинными лучами. Наружная часть прокамбия дифференцируется в первичную флоэму, на периферии которой нередко развиваются механические волокна, внутренняя — в первичную ксилему. Между проводящими тканями остаётся слой клеток, образующих камбий , который откладывает наружу элементы вторичной флоэмы — луба, внутрь — вторичной ксилемы — древесины, обусловливая утолщение стелы.
Строение С. в зоне узлов отличается от строения средней части междоузлия наличием листовых и веточных лакун (прорывов). У двудольных (см. рис. 1) часты трёхлакунные узлы (яблоня), реже встречаются однолакунные (сирень) и многолакунные (бузина). Наиболее активное вторичное утолщение свойственно многолетним древесным растениям, во вторичной древесине которых (а иногда и в лубе) можно видеть границы годичных приростов. С возрастом вследствие развития перидерм первичная кора, а позднее и наружная часть луба отмирают, образуя корку . Для большинства однодольных (см. рис. 1, V) характерен пальмовый тип прохождения закрытых коллатеральных пучков, обусловливающих их диффузное расположение на поперечных срезах. Лишь у некоторых злаков со С.-соломиной, у традесканции и диоскореи имеется тенденция к круговому расположению пучков. Вторичное утолщение свойственно только древовидным лилейным (алоэ, драцена), у которых в перицикле или первичной коре формируется меристема, образующая концентрические пучки и межпучковую (часто одревесневающую) паренхиму. См. также Стелярная теория .
Лит.: Серебряков И. Г., Морфология вегетативных органов высших растений, М., 1952; Мейер К. И., Морфогения высших растений, М., 1958; Имс А., Морфология цветковых растений, пер. с англ., М., 1964; Ботаника, под ред. Л. В. Кудряшова, т. 1, М., 1966; Эсау К., Анатомия растений, пер. с англ., М., 1969.
Л. И. Лотова.
Рис. 1. Анатомическое строение стебля цветковых растений: I — общий вид проводящей системы стебля с причленившимся трехпучковым листовым следом; II — строение стебля в области трехлакунного узла, III — однолакунного, IV — многолакунного; V — пальмовый тип прохождения пучков в стебле однодольных растений; VI — строение соломины злаков; VII — строение стебля бигнонии с вдающимися в древисину участками луба; VIII — строение стебля вистарии, утолщение которого обусловлено несколькими камбиями; 1 — сердцевина; 2 — стела; 3 — листовой след; 4 — пучки листового следа; 5 — листовые прорывы; 6 — флоэмные волокна; 7 — ксилема; 8 — камбий пучковый; 9 — флоэма; 10 — стебель; 11 — влагалище листа; 12 — закрытые коллатеральные пучки; 13 — ассимиляционная паренхима; 14 — воздушная полость; 15 — сосуды ксилемы; 16 — механическая ткань; 17 — сердцевинные лучи; 18 — перидерма; 19 — первичная кора; 20 — древесина; 21 — луб.