Лит.: Кизюрин А. Д., Кустовидно-стелющийся метод сибирского плодоводства, [Омск], 1963.
Е. В. Колесников.
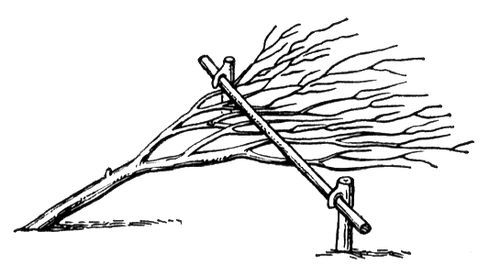
Рис. 1. Минусинский стланец.
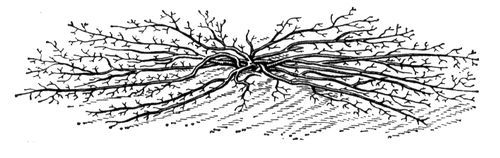
Рис. 2. Бесштамбовый стланец.
Стелющиеся растения
Сте'лющиеся расте'ния, растения с горизонтальными приземными побегами, которые полегают и могут укореняться в процессе роста (самые молодые участки побегов и их верхушечные почки могут быть направлены вверх). Стелющиеся деревья обычно называют стланцами (горная сосна, кедровый стланик, арча), кустарники — стланиками, кустарнички — стланичками (полярные ивы, некоторые виды рододендронов, толокнянка), травянистые С. р. с укоренёнными побегами — ползучими, с неукоренёнными — лежачими. Древесные С. р. приспособлены к суровым почвенно-климатическим условиям и приурочены главным образом к субарктическим, субантарктическим и высокогорным областям. Приземное положение даёт им преимущество в тепловом режиме, водоснабжении, защищает от иссушающих ветров. Стелющиеся стволы и ветви более долговечны, чем прямостоячие. Травянистые С. р. обычно связаны с затенёнными или избыточно увлажнёнными местообитаниями (например, лесные травы — копытень, вероника лекарственная; луговые, болотные и прибрежные — полевица, луговой чай, лютик ползучий).
Т. И. Серебрякова.
Стелярная теория
Стеля'рная тео'рия, учение о принципах строения и взаимоотношениях между типами стелы (центрального цилиндра ) у высших растений. Большую часть стелы составляют проводящие ткани : ксилема (древесина) и флоэма (луб), по-разному расположенные в разных типах стелы. Проводящие ткани обычно окружены перициклом , состоящим из механических и паренхимных клеток. Вокруг стелы находится первичная кора.
Учение о стеле создано французскими ботаниками Ф. Ван Тигемом и А. Дулио (1886), которым принадлежит и первая классификация типов стелы. Дальнейшее развитие С. т. получила в работах английского ботаника Г. Бребнера (1902), американского — Э. Джефри (1903, 1917), значительно усовершенствовавших классификацию типов стелы, а затем в трудах немецкого учёного В. Циммерманна и советских — К. И. Мейера, А. Л. Тахтаджяна, отметивших изменения стелы в онтогенезе и филогенезе растений.
Исходным типом стелы, характерным для древнейших высших растений — псилофитов (риния), считают протостелу, имеющую вид центр, тяжа, во внутренней части которого расположена ксилема, окруженная нерезко отграниченной от первичной коры флоэмой. Совершенствование структуры стелы в эволюции растений, по мнению английского ботаника Ф. Боуэра, шло по пути создания наиболее рационального соотношения между объёмом и поверхностью проводящих тканей, что достигалось изменениями контуров стелы и привело к расчленению её на отдельные тяжи. Большую роль в эволюции стелы сыграли процессы медулляции, т. е. формирования сердцевины, и витализации проводящих тканей — появления в них живых паренхимных клеток. Развитие стелярной структуры растений сопровождалось также дифференциацией прокамбия на протоксилему и метаксилему, протофлоэму и метафлоэму, изменениями характера заложения ксилемы от мезархного к экзархному и эндархному. При мезархном заложении первые элементы ксилемы дифференцируются в центре прокамбиального тяжа, все последующие — в радиальных направлениях. Для экзархной ксилемы характерно периферическое заложение первых элементов и центростремительное развитие последующих. В эндархной ксилеме первые элементы формируются из внутренней части прокамбия, а следующие развиваются центробежно. Большое значение имели расчленение стелы на стеблевые и листовые пучки, входящие в стебель из листа, усиление связи между ними, формирование эндодермы как барьера, предотвращающего потерю влаги и задерживающего в стеле продукты ассимиляции. В процессе исторического развития наземных растений происходило, как правило, увеличение размеров стелы.
В разных направлениях эволюции растений возникали различные типы структурной организации стелы. Так, изменения контуров ксилемы обусловили преобразование протостелы в актиностелу и плектостелу. Актиностела, характерная для псилофитов (астероксилон), а из современных растений — для псилота, имеет лопастные очертания поперечного сечения экзархной ксилемы.
В плектостеле, обычной для плаунов, экзархная ксилема рассечена на лентовидные тяжи. Система проводящих тканей, образующих трубчатый цилиндр, окружающий паренхимную сердцевину, характерна для сифоностелы. У папоротников сифоностела бывает трёх типов: эктофлойная (соленоксилия), амфифлойная (соленостела) и диктиостела. Соленоксилия образовалась, по-видимому, из актиностелы при втягивании внутрь отрогов ксилемы, срастании участков флоэмы в сплошное кольцо и развитии паренхимной сердцевины, клетки которой возникли из трахеид, утративших способность проводить воду и поделившихся поперечными перегородками. В соленоксилии ксилема снаружи окружена флоэмой, перициклом и эндодермой (например, у хельминтостахиса). В соленостеле имеются не только наружные, но и внутренние флоэма, перицикл и эндодерма (например, у марсилии). Исследования К. И. Мейера по развитию проводящей системы папоротников показали возможность образования соленостелы из соленоксилии. В результате сильного рассечения амфифлойной сифоностелы в связи с появлением многочисленных листовых прорывов, заполненных паренхимой, возникла диктиостела, имеющая вид сетчатого цилиндра, т.к. составляющие её проводящие ткани образуют переплетающиеся тяжи (меристелы). На поперечном срезе стебля меристелы расположены кольцом вокруг сердцевины. Они построены по типу концентрических амфикрибральных пучков, в которых флоэма расположена вокруг ксилемы; флоэма окружена перициклом и эндодермой.
Возрастные изменения стелы у папоротников (мараттия, орляк, матония), выражающиеся в формировании внутри одной стелы второй, а затем и третьей, привели к полициклии. Переход от протостелы к полистелии обусловил, по мнению. В. Циммерманна, формирование эустелы, в которой каждая протостела превратилась в коллатеральный пучок. Некоторые ботаники считают, что эустела могла сформироваться из эктофлойной сифоностелы, расчленение которой на отдельные пучки было вызвано образованием сердцевинных лучей. У хвощей эустела представлена закрытыми коллатеральными пучками, расположенными вокруг центральной воздушной полости и соединяющимися в узлах; на месте рано разрушающейся ксилемы в пучке образуется каринальный канал (водоносная полость). Эту разновидность стелы называют артростелой У разных видов хвощей артростелы характеризуются различным расположением эндодермы (см. рис., позиции и, к, л). Исследования ранних этапов развития проводящей системы хвощей дают возможность предполагать, что артростела образовалась из актиностелы или из сифоностелы вследствие расщепления их на отдельные пучки. Для эустелы семенных растений характерно усиление роли листовых пучков и наличие тесных контактов между стеблевыми и листовыми пучками. Эустела двудольных растений представлена системой открытых коллатеральных или биколлатеральных пучков с эндархной первичной ксилемой; пучки разделены паренхимными сердцевинными лучами, пересекающими стелу в радиальном направлении. У многих травянистых растений сердцевинные лучи широкие; у древесных — узкие, иногда однорядные, перицикла и эндодермы нет.
Мощное развитие проводящих пучков листьев, которые, войдя в стебель, располагаются по всему поперечному сечению стебля, а также редукция стеблевых пучков привели к преобразованию эустелы в атактостелу однодольных растений, утратившую способность к вторичному утолщению. Проводящие пучки в атактостеле коллатеральные или концентрические амфивазальные (ксилема в них окружает флоэму).