Внутри каждой парцеллы создается свой фитоклимат. Весной в тенистых еловых парцеллах снег лежит дольше, чем на участках под листопадными деревьями или в окнах. Поэтому активная жизнь весной в парцеллах наступает в разные сроки, переработка детрита также идет с разной скоростью. Границы между парцеллами могут быть как относительно четкими, так и размытыми. Взаимосвязь осуществляется как в результате кондиционирования условий среды (теплообмен, изменение освещения, перераспределение осадков и т. п.), так и в результате материально-энергетического обмена. Происходит разброс растительного опада, перенос пыльцы, спор, семян и плодов воздушными потоками и животными, перемещение животных, поверхностный сток осадков и талых вод, передвигающих минеральные и органические вещества. Все это поддерживает биогеоценоз как единую, внутренне разнородную экосистему.
Роль разных парцелл в строении и функционировании биогеоценозов неодинакова, наиболее крупные парцеллы, занимающие большие пространства и объем, называют основными. Их бывает немного. Именно они определяют внешний облик и строй биогеоценоза. Парцеллы, занимающие небольшие площади, называют дополняющими. Число их всегда больше. Одни парцеллы более устойчивы, другие подвержены значительным и быстрым изменениям. По мере взросления и старения растений парцеллы могут сильно изменить состав и структуру, ритмы сезонного развития, по-разному участвовать в круговороте веществ.
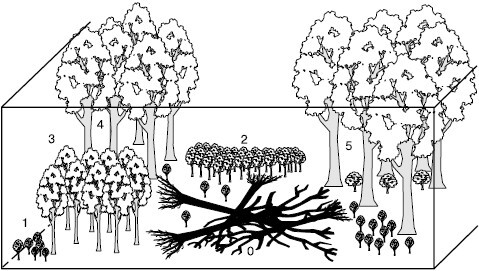
Рис. 145. Окна возобновления основных пород в лесном биогеоценозе (по О. В. Смирновой, 1998)
Мозаичность лесных биогеоценозов и появление новых парцелл часто связаны с образованием в лесах окон, т. е. нарушением древесного яруса в связи с вывалом старых деревьев, вспышек массовых вредителей – насекомых, поражением грибами, деятельностью крупных копытных. Создание такой мозаичности совершенно необходимо для устойчивого существования леса и возобновления главенствующих пород деревьев, подрост которых часто не может развиваться под материнскими кронами, так как требует иных условий освещения и минерального питания. Окна возобновления для разных пород должны иметь достаточную пространственную протяженность (рис. 145). В восточноевропейских широколиственных лесах ни один вид не может переходить к плодоношению в окнах, соизмеримых всего с проекциями крон одного-двух взрослых деревьев. Даже наиболее теневыносливым из них – букам, кленам – требуются освещенные парцеллы в 400–600 м2, а полный онтогенез светолюбивых видов – дуба, ясеня, осины может завершаться только в крупных окнах не менее 1500–2000 м2.
На основании детального изучения структуры и функционирования биогеоценозов в экологии в последнее время развивается концепция мозаично-циклической организации экосистем. С этой точки зрения устойчивое существование многих видов в экосистеме достигается за счет постоянно происходящих в ней естественных нарушений местообитаний, позволяющих новым поколениям занимать вновь освободившееся пространство.
Биогеоценология рассматривает поверхность Земли как сеть соседствующих биогеоценозов, связанных между собой через миграцию веществ, но тем не менее, хотя и в разной степени, автономных и специфичных по своим круговоротам. Конкретные свойства участка, занятого биогеоценозом, придают ему своеобразие, выделяя из других, исходных по типу.
Обе концепции – экосистем и биогеоценозов – дополняют и обогащают друг друга, позволяя рассматривать функциональные связи сообществ и окружающей их неорганической среды в разных аспектах и с разных точек зрения.
9.2. Поток энергии в экосистемах
Поддержание жизнедеятельности организмов и круговорот вещества в экосистемах возможны только за счет постоянного притока энергии (рис. 146). В конечном счете вся жизнь на Земле существует за счет энергии солнечного излучения, которая переводится фотосинтезирующими организмами в химические связи органических соединений. 4етеротрофы получают энергию с пищей. Все живые существа являются объектами питания других, т. е. связаны между собой энергетическими отношениями. Пищевые связи в сообществах – это механизмы передачи энергии от одного организма к другому. В каждом сообществе трофические связи переплетены в сложную сеть. Организмы любого вида являются потенциальной пищей многих других видов. Врагами тлей, например, служат личинки и жуки божьих коровок, личинки мух-сирфид, пауки, насекомоядные птицы и многие другие. За счет дубов в широколиственных лесах могут жить несколько сотен форм различных членистоногих, фитонематод, паразитических грибков и т. п. Хищники обычно легко переключаются с одного вида жертв на другой, а многие, кроме животной пищи, способны потреблять в некотором количестве и растительную. Таким образом, трофические сети в биоценозах очень сложные и создается впечатление, что энергия, поступившая в них, может долго мигрировать от одного организма к другому.
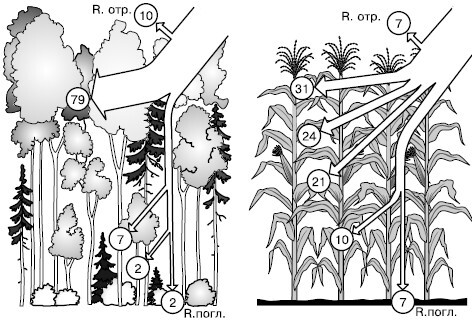
Рис. 146. Распределение радиации (в кружочках – %) в бореальном смешанном лесу и в посевах кукурузы (по В. Лархеру, 1978): R. отр. – радиация, отраженная от поверхности насаждения; R. погл. – радиация, поглощенная почвой
На самом деле путь каждой конкретной порции энергии, накопленной зелеными растениями, короток. Она может передаваться не более чем через 4–6 звеньев ряда, состоящего из последовательно питающихся друг другом организмов. Такие ряды, в которых можно проследить пути расходования изначальной дозы энергии, называют цепями питания (рис. 147).
Место каждого звена в цепи питания называют трофическим уровнем. Первый трофический уровень – это всегда продуценты, создатели органической массы; растительноядные консументы относятся ко второму трофическому уровню; плотоядные, живущие за счет растительноядных форм, – к третьему; потребляющие других плотоядных – соответственно к четвертому и т. д. Таким образом, различают консументов первого, второго и третьего порядков, занимающих разные уровни в цепях питания. Естественно, что основную роль при этом играет пищевая специализация консументов. Виды с широким спектром питания могут включаться в пищевые цепи на разных трофических уровнях. Так, например, человек, в рацион которого входит как растительная пища, так и мясо травоядных и плотоядных животных, выступает в разных пищевых цепях в качестве консумента первого, второго и третьего порядков. Виды, специализированные на растительной пище, например тли, зайцеобразные, копытные, всегда являются вторым звеном в цепях питания.
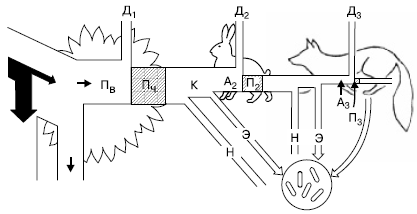
Рис. 147. Поток энергии через три уровня простой пищевой цепи (по П. Дювиньо и М. Тангу, 1968): Пв – продукция валовая; Пч – продукция чистая; К – продукция, использованная на корм; А2, А3 – корм, ассимилированный консументами; н – неиспользованная часть продукции; П2 – вторичная продукция (травоядные); П3 – прирост хищников; Д1-Д3 – траты энергии на обмен веществ (траты на дыхание) на разных уровнях пищевой цепи
Энергетический баланс консументов складывается следующим образом. Поглощенная пища обычно усваивается не полностью. Неусвоенная часть вновь возвращается во внешнюю среду (в виде экскрементов) и в дальнейшем может быть вовлечена в другие цепи питания. Процент усвояемости зависит от состава пищи и набора пищеварительных ферментов организма. У животных усвояемость пищевых материалов варьирует от 12–20 % (некоторые сапрофаги) до 75 % и более (плотоядные виды). Ассимилированная организмом пища вместе с запасом в ней энергии расходуется двояким образом. Большая часть энергии используется на поддержание рабочих процессов в клетках, а продукты расщепления подлежат удалению из организма в составе экскретов (мочи, пота, выделений различных желез) и углекислого газа, образующегося при дыхании. Энергетические затраты на поддержание всех метаболических процессов условно называют тратой на дыхание, так как общие их масштабы можно оценить, учитывая выделение СО2 организмом. Меньшая часть усвоенной пищи трансформируется в ткани самого организма, т. е. идет на рост или откладывание запасных питательных веществ, увеличение массы тела. Эти отношения сокращенно можно выразить формулой: