Живые организмы не только испытывают воздействия со стороны паразитов и симбионтов, но и энергично реагируют на них. Это сопротивление паразитам получило название активного иммунитета. Полноценные, здоровые особи растений и животных часто обладают действенными защитными приспособлениями, не позволяющими проникать в них патогенным организмам. Например, устойчивость хвойных деревьев к нападению стволовых вредителей (жуков-короедов, личинок златок, усачей и пр.) обеспечивается прежде всего выделением смолы, которая содержит соединения, токсические для этих насекомых. Ослабленные деревья теряют сопротивляемость, их заселяют насекомые, поражают грибы и другие потребители живых тканей. У животных защитной реакцией от вторжения посторонних организмов является выработка гуморального иммунитета – образование в крови хозяина специфических белковых веществ, антител, подавляющих паразитов. Выработка иммунитета стимулируется токсинами паразита и часто предохраняет от повторных заражений.
В ряде случаев организм хозяина отвечает на вторжение паразита разрастанием окружающих его тканей, образованием своеобразной капсулы, изолирующей паразита. Такие образования у животных называют зооцецидиями, а у растений – галлами. Иногда такая изоляция приводит к гибели паразита. Например, личинки глист, проникающие под наружный слой мантии ряда пластинчатожаберных моллюсков, окружаются и замуровываются отложениями перламутра -возникает жемчужина. Чаще, однако, защитная роль зооцецидиев ограничивается локализацией причиняемого Mikiola fagi вреда в определенном участке тела хозяина, а сами паразиты используют их как дополнительные защитные образования. Особенно наглядно это выражено в возникновении галлов у растений (рис. 57). Вызывающие их насекомые, клещи, нематоды и другие паразиты выделяют специальные вещества, стимулирующие преобразование тканей или целых органов растения в галлы с камерой внутри, в которой их обитатель надежно защищен от высыхания и врагов и обеспечен пищей.
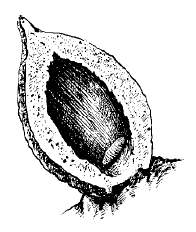
Рис. 57. Разрезанный галл на листе бука с личинкой мушки-галлицы Mikiola fagi
Ограниченность жизненного пространства особенно сказывается на размерах и форме внутриклеточных паразитов и симбионтов. Так, грегарины, живущие в полости кишечника, – это крупные споровики со сложно расчлененной клеткой, тогда как кокцидии или малярийные плазмодии, являющиеся внутриклеточными паразитами, отличаются очень мелкими размерами и упрощенным внешним строением.
Недостаток кислорода в тканях и особенно в желудочно-кишечном тракте организмов-хозяев приводит к тому, что у многоклеточных обитателей внутриорганизменной среды вырабатывается преимущественно анаэробный тип обмена. Необходимая для работы клеток энергия высвобождается не за счет дыхания, а за счет разных видов брожения. У человеческой аскариды утрачены все ферменты дыхательного цикла, и кислород действует на червей как яд, что используется в медицинской практике. Однако целый ряд паразитов не утрачивает полностью способности к дыханию и может переключаться с анаэробного типа обмена на аэробный, например жгутиковые Trichomonas, эхинококк и др.
Среда обитания паразитов ограничена как во времени (жизнью хозяина), так и в пространстве. Поэтому основные адаптации направлены на возможность распространения в этой среде, передачи от одного хозяина к другому. Главнейшие приспособления к этому – повышенная способность к размножению, выработка сложных жизненных циклов, использование переносчиков и промежуточных хозяев.
Громадная плодовитость, свойственная паразитам, получила название «закон большого числа яиц». Естественный отбор привел к развитию у паразитов гигантской яйцепродукции по сравнению со свободноживущими формами. Человеческая аскарида продуцирует в среднем 250 тыс. яиц за сутки, а за всю жизнь – свыше 50 млн. Масса яиц, отложенных одной самкой аскариды за год, в 1700 раз превышает ее собственную массу.
Подавляющее большинство яиц и зародышей паразитов гибнет, не попав в очередного хозяина или не выдержав воздействия различных факторов внешней среды. Только чрезвычайная плодовитость увеличивает шансы на выживание и завершение жизненного цикла хотя бы немногих потомков, поддерживая существование вида.
Приспособления к умножению потомства проявляются у ряда паразитов также в виде партеногенеза, полиэмбрионии (клетки одного делящегося яйца дают начало множеству зародышей), бесполого размножения (почкование у пузырчатых стадий ленточных червей). Это приводит к чередованию поколений – полового и партеногенетического или полового и бесполых.
У многих паразитов чередование поколений сочетается со сменой двух или более хозяев, так что одно поколение существует в одном хозяине, а другое – в другом. Партеногенетическое или бесполое размножение в промежуточном хозяине увеличивает шансы паразита на попадание в окончательного хозяина и завершение жизненного цикла. Многообразие и сложность жизненных циклов паразитов выработались как приспособление для передачи от одной особи хозяина к другой для распространения.
В ряде случаев паразиты сами становятся средой обитания других видов – возникает явление гиперпаразитизма. Двух– или даже трех– и четырехступенчатый паразитизм не так редок в природе. Например, многоядерные опалины, живущие в амфибиях, сами бывают иногда на 100 % заражены амебами, размножающимися внутри их клеток. В свою очередь, амебы, паразитирующие на опалинах, могут быть поражены грибком из рода Sphaerita. Много сверхпаразитов в классе насекомых. Например, для паразита капустной белянки наездника Apanteles glomeratus известно более 20 видов вторичных паразитов из перепончатокрылых.
В разных частях тела многоклеточного организма условия неоднородны. Хозяин выступает для своих обитателей как многообразная среда. Его паразиты и сожители приспособились к жизни в определенных органах и тканях, они приурочены к определенному возрастному и физиологическому состоянию хозяина. Например, в Сибири в лиственницах тонкоусый еловый усач заселяет преимущественно прикомлевую часть до высоты примерно 1 м, лиственничная златка осваивает ствол выше, до 4–5 м, продолговатый короед распространяется по всей его средней части, а вершину и ветви заселяют короед-гравер и заболонник Моравица. В пищеварительном тракте кролика одновременно могут паразитировать несколько видов кокцидий, каждый из которых локализуется в определенных частях кишечника: Eimeria media – в начальном отделе тонкой кишки, Е. irresidua – в средней, а Е. magnа – в последней ее петле, Е. piriformis – преимущественно в слепой кишке и т. д.
Фауна паразитов изменяется с возрастом хозяина. Увеличение его размеров, исчезновение личиночных органов и появление новых, смена сред обитания, метаморфоз, линьки – все это отражается на заселенности хозяина паразитами и симбионтами.
Большой круг паразитов обитает не внутри, а на поверхности тела хозяина. Последний в этом случае выступает лишь как часть внешней среды паразита, снабжая его пищей, предоставляя убежище, трансформируя микроклимат. Связь эктопаразита с хозяином может быть постоянной или временной. Для постоянных или длительно связанных с хозяином эктопаразитов одна из основных жизненно важных экологических задач – удержаться на теле хозяина. В связи с этим типичные эктопаразиты обычно характеризуются наличием мощных органов прикрепления – присосок, крючьев, коготков и т. п., которые независимыми путями развиваются у самых разных по происхождению видов.
Таким образом, паразиты, как и свободноживущие виды, обладают сложной системой приспособлений к своей среде обитания. Их строение и организация отражают специфику этой среды. У представителей разных групп, ведущих паразитический образ жизни, часто развиваются сходные типы приспособлений.
Глава 5. АДАПТИВНЫЕ БИОЛОГИЧЕСКИЕ РИТМЫ
Одно из фундаментальных свойств живой природы – цикличность большинства происходящих в ней процессов. Вся жизнь на Земле, от клетки до биосферы, подчинена определенным ритмам. Природные ритмы для любого организма можно разделить на внутренние (связанные с его собственной жизнедеятельностью) и внешние (циклические изменения в окружающей среде).