Разграничение факторов динамики численности организмов на модифицирующие и регулирующие имеет большое практическое значение из-за принципиальных различий их влияния на популяцию.
Изучение модифицирующих факторов важно для выяснения причин колебания численности и их прогнозирования, изучение регуляторных механизмов – для ограничения амплитуды колебания численности и ее максимально возможной стабилизации.
Таким образом, как масштабы, так и ход колебания численности любого вида в природных сообществах исторически обусловлены естественным отбором в зависимости от особенностей биологии, характера внутривидовых связей и межвидовых отношений.
Как правило, численность видов, имеющих много врагов и конкурентов, ограничивается в биоценозах именно внешними регуляторами, их функциональными и количественными реакциями. Собственные механизмы гомеостаза выявляются лишь при очень высокой плотности популяций, когда возникают сбои в активности потребителей. Виды, которых «все едят», отзываются лишь на чрезмерное увеличение собственной численности, грозящей подрывом ресурсов. Наоборот, крупные хищники и другие виды, испытывающие слабый пресс естественных врагов, обладают очень эффективными внутривидовыми способами регуляции плотности популяции.
8.6.3. Типы динамики численности популяций
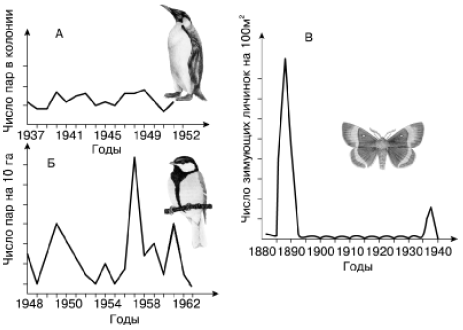
Можно выделить три основных типа популяционной динамики численности (рис. 139).
Рис. 139. Основные типы динамики численности популяций (по М. Уильямсону, 1975): А – пингвина великолепного; Б – большой синицы; В – зимующих гусениц соснового шелкопряда
I. Стабильный тип – отличается небольшим размахом колебаний (в несколько раз, однако не на несколько порядков величин). Свойствен видам с хорошо выраженными механизмами популяционного гомеостаза, высокой выживаемостью, низкой плодовитостью, большой продолжительностью жизни, сложной возрастной структурой, развитой заботой о потомстве. Целый комплекс эффективно работающих регуляторных механизмов держит такие популяции в определенных пределах плотности. Такова, например, динамика численности крупных млекопитающих и птиц, а также ряда беспозвоночных.
II. Флюктуирующий тип – колебания происходят в значительном интервале плотностей, различающихся на один-два порядка величин. При этом различают три фазы колебательного цикла: нарастания, максимума, разрежения численности. Возврат к стабильному состоянию происходит быстро. Регуляторные механизмы не теряют контроля за численностью популяций, увеличивая свою эффективность вслед за увеличением плотности. Преобладают слабоинерционные меж– и внутривидовые взаимодействия. Такой ход численности широко распространен в разных группах животных.
III. Взрывной тип с вспышками массового размножения – прекращение действия модифицирующих факторов не вызывает быстрого возврата популяции в стабильное состояние. Динамика численности складывается из циклов, в которых различают пять обязательных фаз: нарастания численности, максимума, разреживания, депрессии, восстановления. Для популяций периодически характерны предельно высокий и необычайно низкий уровень численности. По фазам цикла также сильно меняются показатели размножения, возрастной и половой структуры популяции, физиологического состояния, поведения, а иногда и морфологических особенностей составляющих ее особей. Такой ход численности обнаруживается чаще всего у видов с малой продолжительностью жизни, высокой плодовитостью, быстрым оборотом генераций. Он свойствен, например, некоторым насекомым (саранчовые, вредители леса – усачи, короеды, ряд чешуекрылых и пилильщиков и др.), среди млекопитающих отмечен у многих видов мышевидных грызунов.
Тип динамики численности – скорее популяционная, но не видовая характеристика. Популяции одних и тех же видов в различных условиях могут характеризоваться разным ходом динамики численности. Это объясняется преимущественно тем, что среди регуляторных механизмов большую роль играют межвидовые взаимосвязи, которые в пределах ареала вида могут быть разной степени напряженности. Так, многие виды, которые в природных условиях сдерживались врагами, проявляют склонность к вспышкам массового размножения в садах и на полях, где ослаблен биологический контроль.
8.6.4. Механизмы динамики численности
Типы популяционной динамики численности наиболее хорошо изучены у лесных насекомых, на примере которых выявляется большое многообразие конкретных связей и особенностей видов, влияющих на плотность их популяций.
В лесных сообществах преобладают виды со стабильным ходом популяционной динамики. Они питаются практически за счет всех частей деревьев (стволы, корни, побеги, листья, почки, плоды, семена), однако при этом не оказывают губительного воздействия на растения. Как правило, такие индифферентные виды отличаются низкой миграционной активностью, ведут оседлый образ жизни. Комплекс их паразитов включает обычно несколько сильно специализированных видов, эффективно поражающих популяцию хозяина.
Флюктуирующий тип динамики численности свойствен многим ксилофагам (потребителям коры и древесины): усачам, златкам, короедам. Для них характерно совместное заселение кормовых объектов – ослабление деревьев. Это позволяет быстро снизить устойчивость дерева, но в то же время совместное обитание ксилофагов обостряет конкурентные отношения между ними, что действует как безынерционный механизм регуляции численности.
Для короедов, поселяющихся первыми на ослабленных деревьях, при избыточном запасе питательных веществ (растворимых углеводов и крахмала) развитие лимитируется воздействием защитных реакций дерева, таких, например, как смоловыделение у хвойных. Кроме того, еще отсутствуют симбиотические микроорганизмы. Сопротивление еще жизнеспособных деревьев может быть сломлено при концентрированном нападении и избыточно высокой плотности вредителя. Другой предел пригодности дерева для короедов – полное отмирание и разрушение луба. Между двумя указанными состояниями дерева для короедов создается оптимум кормовых условий, отмечается максимальная выживаемость всех фаз развития и наиболее высокие коэффициенты размножения.
Относительную роль различных регуляторных механизмов, определяющих флюктуирующий ход численности лесных насекомых, можно проследить на примере большого лиственного короеда – Ips subelongatus. Этот вредитель лиственницы и частично других пород широко распространен по всей таежной зоне.
Жуки нападают на физиологически ослабленные живые лиственницы, у которых отсутствует активное защитное смоловыделение. Внедряясь под кору, самцы продуцируют запаховые вещества – феромоны, привлекающие других особей, чем обеспечивается массовость заселения подходящего дерева и его дальнейшее ослабление. Феромоны играют также роль химических регуляторов внутрипопуляционных отношений – распределения короедов по стволу, своего рода меткой территории отдельных гнезд. Жуки, не нашедшие места на данном дереве, переселяются на другие. В дальнейшем внутривидовая конкуренция регулируется самками, которые в зависимости от плотности поселения изменяют длину маточных ходов и количество откладываемых яиц. При возрастающем недостатке площади часть самцов и самок мигрирует на другие объекты. Личинка короеда за период развития потребляет 0,78 см6 луба. Если «кормовые площадки» личинок при высокой плотности поселения становятся меньше этого предела, среди них увеличивается смертность. Таким образом, внутривидовая конкуренция регулирует заселенность дерева на всех фазах жизни вредителя: на стадии взрослых жуков она ведет к усилению миграций, на стадии личинок – к гибели части популяции. Показатели размножения остаются высокими на всех этапах флюктуации численности популяции.