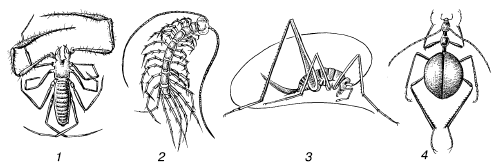
Рис. 12.Ложногомойотермные членистоногие – обитатели пещер: 1– лжескорпион; 2 – многоножка; 3 – кузнечик; 4 – жужелица
Для характеристики организмов по основным источникам используемого тепла используют термины эктотермный и эндотермный. Эктотермия – это жизнь преимущественно за счет нагревания из внешней среды, эндотермия – за счет тепла, вырабатываемого самим организмом.
Масштабы выработки тепла сильно отличаются у разных видов, проявляя зависимость от сложности организации группы, возможностей окислительных реакций, размеров и массы тела, условий среды и других причин. Так, например, бактерии выделяют на грамм веса в час около 450 кал, мухи-дрозофилы – 30, мыши – 8, для человека этот показатель равен 4. В пределах позвоночных животных при сходной массе тела млекопитающие продуцируют в 5–6 раз, а птицы – в 7–8 раз больше тепла, чем рептилии.
Все живые организмы потенциально эндотермны, но сильно различаются по уровню обмена и возможностям сохранения тепла. Нарушения теплового баланса меняют температуру тела. Восстановить нарушенный баланс можно тремя путями: 1) изменением теплопродукции, 2) изменением теплоотдачи и 3) перемещением в пространстве в область предпочитаемых температур. Пойкилотермные и гомойотермные организмы по-разному реализуют возможности температурных адаптаций.
3.1.3. Температурные адаптации пойкилотермных организмов
Температура пойкилотермных изменяется вслед за температурой окружающей среды. Они преимущественно эктотермны, выработки и сохранения собственного тепла у них недостаточно для противостояния тепловому режиму местообитаний. В связи с этим реализуется два основных пути адаптации: специализация и толерантность.
Специализированные виды стенотермны, они приспособлены к жизни в таких участках биосферы, где колебания температур происходят лишь в узких пределах. Выход за эти пределы для них губителен. Например, некоторые одноклеточные водоросли, развивающиеся в горных ледниках на поверхности тающего льда, погибают при температурах, превышающих +(3–5) °С. Растения дождевых тропических лесов не способны переносить снижение температуры до +(5–8) °С. Коралловые полипы живут только в диапазоне температур воды от +20,5 до +30 °C, т. е. в тропическом поясе океана. Голотурия Elpidia glacialis обитает при температуре воды от 0 до +1 °C и не выдерживает отклонения от этого режима ни на один градус.
Другой путь адаптации пойкилотермных видов – развитие устойчивости клеток и тканей к широкому колебанию температур, характерному для большей части биосферы. Этот путь связан с периодическим торможением обмена веществ и перехода организмов в латентное состояние, когда температура среды сильно отклоняется от оптимума.
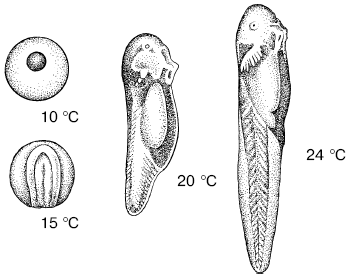
Эффективные температуры развития пойкилотермных организмов. Зависимость темпов роста и развития от внешних температур дает возможность рассчитать прохождение жизненного цикла видов в конкретных условиях. После холодового угнетения нормальный обмен веществ восстанавливается для каждого вида при определенной температуре, которая называется температурным порогом развития, или биологическим нулем развития. Чем больше температура среды превышает пороговую, тем интенсивнее протекает развитие и, следовательно, тем скорее завершается прохождение отдельных стадий и всего жизненного цикла организма (рис. 13).
Рис. 13. Состояние развивающихся при разных температурах головастиков через 3 дня после оплодотворения яйца (по С. А. Зернову, 1949)
Для осуществления генетической программы развития пой-килотермным организмам необходимо получить извне определенное количество тепла. Это тепло измеряется суммой эффективных температур. Под эффективной температурой понимают разницу между температурой среды и температурным порогом развития организмов. Для каждого вида она имеет верхние пределы, так как слишком высокие температуры уже не стимулируют, а тормозят развитие.
И порог развития, и сумма эффективных температур для каждого вида свои. Они зависят от исторической приспособленности к условиям жизни. Для семян растений умеренного климата, например гороха, клевера, порог развития низкий: их прорастание начинается при температуре почвы от 0 до +1 °C; более южные культуры – кукуруза и просо – начинают прорастать только при +(8-10) °С, а семенам финиковой пальмы для начала развития нужно прогревание почвы до +30 °C.
Сумму эффективных температур рассчитывают по формуле
X = (T – C) · t,
где X– сумма эффективных температур; T – температура окружающей среды, С – температура порога развития и t – число часов или дней с температурой, превышающей порог развития.
Зная средний ход температур в каком-либо районе, можно рассчитать появление определенной фазы или число возможных генераций интересующего нас вида. Так, в климатических условиях Северной Украины может выплодиться лишь одна генерация бабочки яблонной плодожорки, а на юге Украины – до трех, что необходимо учитывать при разработке мер защиты садов от вредителей. Сроки цветения растений зависят от того, за какой период они набирают сумму необходимых температур. Для зацветания мать-и-мачехи под Петербургом, например, сумма эффективных температур равна 77, кислицы – 453, земляники – 500, а желтой акации – 700 °C.
Сумма эффективных температур, которую нужно набрать для завершения жизненного цикла, часто ограничивает географическое распространение видов. Например, северная граница лесной растительности приблизительно совпадает с июльскими изотермами +(10–12) °С. Севернее тепла для развития деревьев уже не хватает, и зона лесов сменяется безлесными тундрами.
Расчеты эффективных температур необходимы в практике сельского и лесного хозяйства, при борьбе с вредителями, интродукции новых видов и т. п. Они дают первую, приближенную основу для составления прогнозов. Однако на распространение и развитие организмов влияет множество других факторов, поэтому в действительности температурные зависимости оказываются более сложными.
Температурная компенсация. Ряд пойкилотермных видов, обитающих в условиях переменных температур, развивает возможность поддерживать более или менее постоянный уровень обмена веществ в довольно широких пределах изменения температуры тела. Это явление называется температурной компенсацией и происходит в основном за счет биохимических адаптаций. Например, у моллюсков на побережье Баренцева моря, таких, как брюхоногие литторины (Littorina littorea) и двустворчатые мидии (Mytilus edulis), интенсивность обмена, оцениваемая по потреблению кислорода, почти не зависит от температуры в тех пределах, с которыми моллюски встречаются ежедневно во время приливов и отливов. В весенне-летний период этот диапазон достигает более 20 °C (от +6 до +30 °C), и в холодной воде их метаболизм столь же интенсивен, как в теплом воздухе. Это обеспечивается действием ферментов, которые при понижении температуры меняют свою конфигурацию таким образом, что возрастает их сродство к субстрату и реакции протекают более активно.
Другие способы температурной компенсации связаны с заменой действующих ферментов сходными по функции, но работающими при иной температуре (изоферментами). Такие адаптации требуют времени, поскольку происходит инактивация одних генов и включение других с последующими процессами сборки белков. Подобная акклимация (сдвиг температурного оптимума) лежит в основе сезонных перестроек, а также обнаруживается у представителей широко распространенных видов в разных по климату частях ареала. Например, у одного из видов бычков из Атлантического океана в низких широтах Q10 имеет невысокое значение, а в холодных северных водах возрастает при низких температурах и снижается при средних. Результатом этих компенсаций является то, что животные могут поддерживать относительное постоянство активности, так как даже незначительное повышение температуры у критических точек усиливает обменные процессы. Температурные компенсации для каждого вида возможны лишь в определенном диапазоне температур, но не выше и не ниже этой области.