Оттепели в середине, а особенно в конце зимы вызывают быстрое снижение устойчивости растений к морозам. После окончания зимнего покоя закалка утрачивается. Весенние заморозки, наступившие внезапно, могут повредить тронувшиеся в рост побеги и особенно цветки даже у морозоустойчивых растений.
По степени адаптации к высоким температурам можно выделить следующие группы растений:
1) нежаростойкие растения повреждаются уже при +(30–40) °С (эукариотические водоросли, водные цветковые, наземные мезофиты);
2) жаровыносливые растения переносят получасовое нагревание до +(50–60) °С (растения сухих местообитаний с сильной инсоляцией – степей, пустынь, саванн, сухих субтропиков и т. п.).
Некоторые растения регулярно испытывают влияние пожаров, когда температура кратковременно повышается до сотен градусов. Пожары особенно часты в саваннах, в сухих жестколистных лесах и кустарниковых зарослях типа чапарраля. Там выделяют группу растений-пирофитов, устойчивых к пожарам. У деревьев саванн на стволах толстая корка, пропитанная огнеупорными веществами, надежно защищающими внутренние ткани. Плоды и семена пирофитов имеют толстые, часто одревесневшие покровы, которые растрескиваются, будучи опалены огнем.
Возможности регуляции температуры у пойкилотермных животных. Важнейшая особенность животных – их подвижность, способность перемещаться в пространстве создает принципиально новые адаптивные возможности, в том числе и в терморегуляции. Животные активно выбирают местообитания с более благоприятными условиями.
В отличие от растений, животные, обладающие мускулатурой, производят гораздо больше собственного, внутреннего тепла. При сокращении мышц освобождается значительно больше тепловой энергии, чем при функционировании любых других органов и тканей, так как КПД использования химической энергии для совершения мышечной работы относительно низок. Чем мощнее и активнее мускулатура, тем больше тепла может генерировать животное. По сравнению с растениями животные обладают более разнообразными возможностями регулировать, постоянно или временно, температуру собственного тела.
Пойкилотермные животные остаются, однако, как и растения, эктотермными, поскольку общий уровень их метаболизма не настолько высок, чтобы внутреннего тепла стало достаточно для обогревания тела. Например, при температуре +37 °C пустынная игуана потребляет кислорода в 7 раз меньше, чем грызуны такой же величины. Тем не менее некоторые из пойкилотермных животных в состоянии активности способны поддерживать температуру тела более высокую, чем в окружающей среде. Например, бабочки-бражники, ведущие ночной образ жизни, летают и кормятся на цветках даже при +10 °C. Во время полета температура грудного отдела поддерживается на уровне 40–41 °C. Другие насекомые могут летать в холодном воздухе, предварительно разогревая свои летательные мышцы для взлета, например: саранча, шмели, осы, пчелы, крупные ночные совки и др. Шмели собирают нектар даже при +5 °C, имея температуру тела 36–38 °C. При прекращении активности насекомые быстро остывают. Генерировать тепло для обогревания могут в некоторых случаях и рептилии. Самка питона, обвивающая своим телом кладку, сокращая мускулатуру, способна повышать температуру на 5–6 °C в диапазоне внешних температур от +25 до +33 °C. При этом потребление ею кислорода возрастает почти в 10 раз до предельного для рептилий уровня. В более прохладном воздухе змея становится вялой и неактивной.
Основные способы регуляции температуры тела у пойкилотермных животных – поведенческие: перемена позы, активный поиск благоприятных мест обитания, целый ряд специализированных форм поведения, направленных на создание микроклимата (рытье нор, сооружение гнезд и др.).
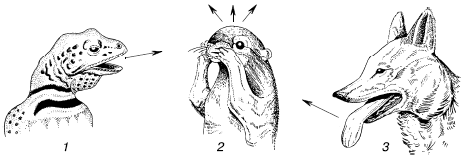
Переменой позы животное может усилить или ослабить нагревание за счет солнечной радиации. Например, пустынная саранча в прохладные утренние часы подставляет солнечным лучам широкую боковую поверхность тела, а в полдень – узкую спинную. Ящерицы даже высоко в горах в период нормальной активности могут поддерживать температуру тела, используя нагревание прямыми солнечными лучами и тепло нагретых скал. По исследованиям на Кавказе, на высоте 4100 м температура тела Lacerta agilis временами на 29 °C превышала температуру воздуха, держась на уровне 32–36 °C. В сильную жару животные прячутся в тень, скрываются в норах, щелях и т. п. В пустынях днем, например, некоторые виды ящериц и змей взбираются на кусты или зарываются в менее нагретые слои песка, избегая соприкосновения с раскаленной поверхностью грунта. Ящерицы при необходимости стремительно перебегают горячие поверхности только на задних ногах, уменьшая тем самым контакт с почвой (рис. 16). К зиме многие животные ищут убежища, где ход температур более сглажен по сравнению с открытыми местами обитания. Еще более сложны формы поведения общественных насекомых: пчел, муравьев, термитов, которые строят гнезда с хорошо регулируемой внутри них температурой, почти постоянной в период их активности.
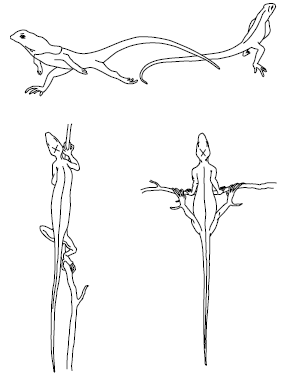
Рис. 16. Поведение ящериц, спасающихся от раскаленной поверхности песка в пустыне
Рис. 17. Испарительная терморегуляция у животных:
1– ящерица – испарение со слизистых при открытом рте;
2– антилоповый суслик – натирание слюной;
3– койот – испарение со слизистых при учащенном дыхании
У ряда пойкилотермных животных эффективно действует и механизм испарительной терморегуляции. Лягушка за час при +20 °C теряет на суше 7770 Дж, что в 300 раз больше ее собственной теплопродукции. Многие рептилии при приближении температуры к верхней критической начинают тяжело дышать или держать рот открытым, усиливая отдачу воды со слизистых оболочек (рис. 17). Пчелы, летающие в жаркую погоду, избегают перегрева, выделяя изо рта каплю жидкости, испарение которой удаляет избыток тепла.
Однако, несмотря на ряд возможностей физической и поведенческой терморегуляции, пойкилотермные животные могут осуществлять ее лишь в узком диапазоне температур. Из-за общего низкого уровня метаболизма они не могут обеспечить постоянство теплового баланса и достаточно активны только вблизи от верхних температурных границ существования. Овладение местообитаниями с постоянно низкими температурами для холоднокровных животных затруднительно. Оно возможно только при развитии специализированной криофилии и в наземных условиях доступно лишь мелким формам, способным использовать малейшие преимущества микроклимата.
3.1.4. Температурные адаптации гомойотермных организмов
Гомойотермия – принципиально иной путь температурных адаптаций, возникший на основе резкого повышения уровня окислительных процессов у птиц и млекопитающих в результате эволюционного совершенствования кровеносной, дыхательной и других систем органов. Потребление кислорода на 1 г массы тела у теплокровных животных в десятки и сотни раз больше, чем у пойкилотермных.
Основные отличия гомойотермных животных от пойкилотермных организмов: 1) мощный поток внутреннего, эндогенного тепла; 2) развитие целостной системы эффективно работающих терморегуляторных механизмов, и в результате 3) постоянное протекание всех физиологических процессов в оптимальном температурном режиме.
Гомойотермные сохраняют постоянный тепловой баланс между теплопродукцией и теплоотдачей и соответственно поддерживают постоянную высокую температуру тела. Организм теплокровного животного не может быть временно «приостановлен» так, как это происходит при гипобиозе или криптобиозе у пойкилотермных.
Гомойотермные животные всегда вырабатывают определенный минимум теплопродукции, обеспечивающий работу кровеносной системы, органов дыхания, выделения и других, даже находясь в покое. Этот минимум получил название базального метаболизма. Переход к активности усиливает выработку тепла и соответственно требует усиления теплоотдачи.