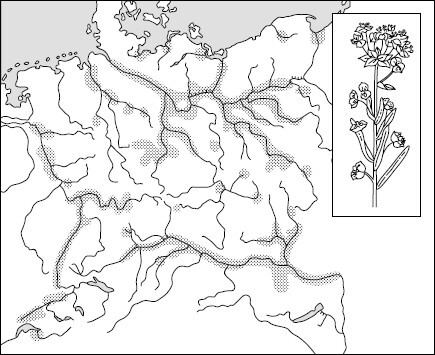
Рис. 95. Ленточные популяции болотного молочая в Средней Европе (по А. Г. Воронову, 1963)
Между популяциями осуществляется обмен отдельными особями, который может быть либо достаточно регулярным, либо эпизодическим. При сезонных кочевках ворон, например, часть молодых птиц ежегодно остается на местах зимовок, образуя пары с представителями оседлого населения. Связь между населением отдельных видов рыб в озерах осуществляется значительно реже, например в годы с особо сильными паводками, когда обособленные водоемы соединяются в единую водную систему.
Взаимоотношения видов не сводятся только к взаимодействию особей, хотя и осуществляются через индивидуальные контакты. Отношения между хищником и его жертвой кратко-временны. Связь между их популяциями длительна и является одним из условий функционирования сообществ. Связи между отдельными особями приводят к одним результатам, связи между популяциями – к качественно другим. Например, длительное воздействие конкретного паразита может иметь следствием изменение физиологического состояния, плодовитости, долговечности хозяина и т. п. Взаимоотношения популяций этих же видов приводят к изменениям их основных групповых характеристик: численности, возрастного состава, смертности, темпов роста популяций.
Связи между популяциями поддерживают вид как единое целое. Слишком длительная и полная изоляция популяций приводит обычно к образованию новых видов.
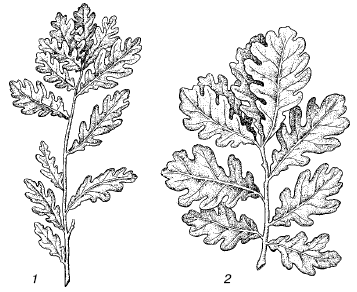
Различия между отдельными популяциями выражены в разной степени. Они могут затрагивать не только их групповые характеристики, но и качественные особенности физиологии, морфологии и поведения отдельных особей (рис. 96). Эти различия создаются в основном под влиянием естественного отбора, приспосабливающего каждую популяцию к конкретным условиям ее существования.
Рис. 96. Ветви дубов, выращенных из желудей, собранных из разных географических популяций: 1– в байрачных лесах Волгоградской области; 2 – в дубравах Белоруссии (по М. С. Двораковскому, 1983)
Зайцы-беляки из разных частей ареала различаются характером окраски, размерами, строением пищеварительной системы. Например, длина слепой кишки у зайцев полуострова Ямал в 2 раза больше, чем у представителей из лесостепного Урала. Это связано с характером питания, разной долей грубых кормов в составе рациона. Средняя величина выводка в центрально-якутской популяции составляет около 7 зайчат на самку, которая дает приплод один раз в год. В карельской части ареала зайчихи приносят зайчат дважды за сезон, но средняя величина выводка около 4 экземпляров. В Белоруссии самки беляков могут давать до четырех приплодов в год. Заячье население разных географических территорий испытывает неодинаковое давление со стороны хищников и паразитов. Например, в Верхоянье до 70 % зайцев бывает поражено цестодами, вызывающими заболевание мозга. Архангельская популяция больше других страдает от печеночного сосальщика. Различия в характере размножения и масштабах гибели от тех или иных причин определяют различия в общей численности, плотности, темпах роста и структуре популяций разных регионов. Популяции различаются между собой тем сильнее, чем более несходны условия их обитания и чем слабее между ними обмен особями.
8.2.2. Классификация популяций
Экологи руководствуются различными принципами для выделения и классификации популяций как территориальных группировок в пределах вида.
Н. П. Наумов на примере млекопитающих рассматривает вид как иерархическую систему популяций различных рангов. Его классификация основывается на ландшафтно-биотопическом подходе к выделению популяционных единиц. Наиболее крупные территориальные группировки вида – подвиды, или географические расы. Система подвидов и масштабы занимаемой ими территории зависят от биологических особенностей вида. Ареалы подвидов у подвижных форм могут быть очень велики. В их пределах на территориях с однородными географическими условиями выделяются географические популяции, отличающиеся общностью приспособлений к климату и ландшафту. Они, в свою очередь, состоят из более мелких популяций, населяющих различные участки среды. К популяциям низшего ранга применяют разные наименования: экологические, биотопические, местные, локальные, элементарные и т. д. Такие популяции могут быть временными и нестабильными. Чем ниже ранг популяций, тем более тесна связь между соседними популяциями, больше степень обмена особями, менее выражены отличительные особенности. Наиболее сильны различия между группировками подвидового ранга. Они затрагивают не только физиологические и поведенческие черты отдельных особей, но и наследственно закреплены в некоторых чертах их морфологии. Связи между популяциями различных рангов обеспечивают единство вида и обогащение его наследственного фонда.
В школе академика С. С. Шварца развивается другой, историко-генетический подход к выделению природных популяций. С этой точки зрения популяции как генетическое единство можно выделять только у видов с половым размножением и перекрестным оплодотворением. Обязательным признаком популяции считается также ее способность к самостоятельному существованию на данной территории в течение неопределенно долгого времени за счет размножения, а не притока особей извне. Временные поселения разных масштабов не относятся к разряду популяций, а считаются внутрипопуляционными подразделениями. С этих позиций вид представлен не иерархическим соподчинением, а пространственной системой соседствующих популяций разных масштабов и с разной степенью связей и изоляции между ними.
С позиций В. Н. Беклемишева и его последователей популяционная структура характерна для всех видов, но при этом следует выделять разные типы популяций, используя критерии, отражающие разные стороны их взаимодействия со средой. Например, по способу размножения и степени генетической целостности популяции делятся на панмиктические (с перекрестным оплодотворением, наиболее целостные), клональные и клонально-панмиктические. Примеры популяций последнего типа можно выделить у тлей, где партеногенетические поколения сменяются половыми. По способности к самовоспроизведению различают постоянные и временные популяции. Постоянные популяции могут быть независимыми, не нуждающимися в притоке особей извне для поддержания своей численности, и полузависимыми, когда такой приток существенно повышает численность популяции, но она может сохраняться и без него. Временные популяции могут также быть различного типа. Общие их особенности в том, что собственный приплод не покрывает смертность и длительность их существования зависит от мигрантов. По размерам различают карликовые, обычные локальные и суперпопуляции. Последние занимают сплошь обширные территории и состоят из большого числа особей. Единство таких популяций в основном топографическое, так как взаимные контакты особей ослаблены расстоянием. В их пределах вычленяют субпопуляции разных масштабов.
Популяции можно классифицировать также по их пространственной и возрастной структуре, по постоянству приуроченности или смене сред обитания и другим экологическим критериям.
Территориальные границы популяций разных видов не совпадают. Наряду с видами, у которых более или менее отчетливо вычленяются локальные популяции (мышевидные грызуны и другие мелкие млекопитающие, некоторые немигрирующие насекомые, моллюски и т. п.), существуют виды с более широкими популяционными границами. Например, территория, занимаемая одной популяцией таких крупных подвижных животных, как лоси, включает множество конкретных участков с разнообразной растительностью: леса с господством различных пород, поля, луга, овраги, поймы рек и т. п. Не ограничены местными пределами популяции многих птиц, крупных млекопитающих, а также ряда мелких организмов с эффективными способами расселения.