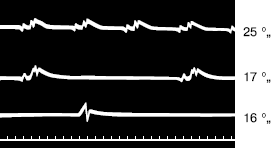
Рис. 22. Изменение числа сердцебиений у сурка вовремя спячки при температуре тела (сверху вниз) 25, 17 и 16 °C. Одно деление шкалы равно 0,2 с (по Эндрес и др., 1930)
Торпидное состояние, или оцепенелость, сопровождает зимнюю спячку животных, но возникает и в других условиях как самостоятельная адаптация. Ряд мелких видов птиц и млекопитающих понижает уровень обмена и температуру тела при неблагоприятных погодных изменениях или регулярно в суточных циклах. У колибри, например, с наступлением ночи температура тела падает с 36–40 °C до 18 °C, птицы садятся на ветки и оцепеневают. Летучие мыши, наоборот, активны в сумеречное и ночное время и впадают в торпидное состояние днем. Переход к оцепенению стимулируется также недостатком пищи. При одних и тех же температурных условиях обеспеченные кормом животные могут продолжать активный образ жизни и генерировать тепло на терморегуляцию, а при нехватке пищи оцепеневают. Кратковременное торпидное состояние, как и спячка, – также регулируемый процесс, но температура тела при этом не падает столь низко и обычно остается на уровне, близком к 20 °C.
Между кратковременным оцепенением гетеротермных животных и длительной зимней спячкой существует вся гамма переходов. Зимний сон медведей, например, не называют спячкой, так как температура этих зверей понижается всего на 3–6 °C, а снижение уровня метаболизма не столь значительно. Мышечная активность обычно подавлена, но большинство физиологических функций сохраняется на прежнем уровне. Роды и выкармливание медвежат молоком у медведиц, например, приходятся на время зимнего сна. Медведи способны относительно легко пробуждаться и переходить в активное состояние.
В пустынях ряд мелких млекопитающих (мышевидные грызуны, сони, некоторые суслики, насекомоядные тенреки и др.) впадают не только в зимнюю, но и в летнюю спячку, которая называется эстивацией (от лат. aestes– лето). Температура их тела падает при этом не столь значительно – до 25–27 °C, но соответственное понижение уровня обмена веществ позволяет экономить не только энергию, но и воду.
Таким образом, широкое распространение гетеротермности позволяет предполагать фундаментальное значение этого явления для выживания животных в неблагоприятных условия.
3.2. Свет
3.2.1. Солнечная радиация
Всем живым организмам для осуществления процессов жизнедеятельности необходима энергия, поступающая извне. Основным источником ее является солнечная радиация, на которую приходится около 99,9 % в общем балансе энергии Земли.
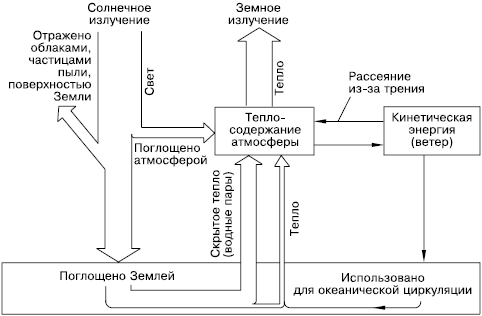
Если принять солнечную энергию, достигающую Земли, за 100 %, то примерно 19 % ее поглощается при прохождении через атмосферу, 34 % отражается обратно в космическое пространство и 47 % достигает земной поверхности в виде прямой и рассеянной радиации (рис. 23). Прямая солнечная радиация – это континуум электромагнитного излучения с длинами волн от 0,1 до 30000 нм. На ультрафиолетовую часть спектра приходится от 1 до 5 %, на видимую – от 16 до 45 % и на инфракрасную – от 49 до 84 %.
Рис. 23. Пути расходования солнечной энергии на поверхности Земли (по Э. Оорту, 1972)
Распределение энергии по спектру существенно зависит от массы атмосферы и меняется при различных высотах Солнца. Количество рассеянной радиации (отраженные лучи) возрастает с уменьшением высоты стояния Солнца и увеличением мутности атмосферы. Спектральный состав радиации безоблачного неба характеризуется максимумом энергии в 400–480 нм.
Действие разных участков спектра солнечного излучения на живые организмы. Среди ультрафиолетовых лучей (УФЛ) до поверхности Земли доходят только длинноволновые (290–380 нм), а коротковолновые, губительные для всего живого, практически полностью поглощаются на высоте около 20–25 км озоновым экраном – тонким слоем атмосферы, содержащим молекулы О3. Длинноволновые ультрафиолетовые лучи, обладающие большой энергией фотонов, имеют высокую химическую активность. Большие дозы их вредны для организмов, а небольшие необходимы многим видам. В диапазоне 250–300 нм УФЛ оказывают мощное бактерицидное действие и у животных вызывают образование из стеролов антирахитичного витамина D; при длине волны 200–400 нм вызывают у человека загар, который является защитной реакцией кожи. Инфракрасные лучи с длиной волны более 750 нм оказывают тепловое действие.
Видимая радиация несет приблизительно 50 % суммарной энергии. С областью видимой радиации, воспринимаемой человеческим глазом, почти совпадает ФР – физиологическая радиация (длина волны 300–800 нм), в пределах которой выделяют ФАР – область фотосинтетически активной радиации (380–710 нм). Область ФР можно условно разделить на ряд зон: ультрафиолетовую (менее 400 нм), сине-фиолетовую (400–500 нм), желто-зеленую (500–600 нм), оранжево-красную (600–700 нм) и дальнюю красную (более 700 нм).
Видимый свет для фототрофных и гетеротрофных организмов имеет разное экологическое значение.
Зеленым растениям свет нужен для образования хлорофилла, формирования гранальной структуры хлоропластов; он регулирует работу устьичного аппарата, влияет на газообмен и транспирацию, активизирует ряд ферментов, стимулирует биосинтез белков и нуклеиновых кислот. Свет влияет на деление и растяжение клеток, ростовые процессы и на развитие растений, определяет сроки цветения и плодоношения, оказывает формообразующее воздействие. Но самое большое значение имеет свет в осуществлении процесса фотосинтеза. С этим связаны основные адаптации растений по отношению к свету.
Фотоавтотрофы способны ассимилировать СО2, используя лучистую энергию Солнца и преобразуя ее в энергию химических связей в органических соединениях. Пурпурные и зеленые бактерии, имеющие бактериохлорофиллы, способны поглощать свет в длинноволновой части (максимумы в области 800-1100 нм). Это позволяет им существовать даже при наличии только невидимых инфракрасных лучей. Водоросли и высшие зеленые растения поглощают свет в диапазоне, близком к видимому человеческим глазом.
Водоросли обитают в водоемах, но встречаются и на суше на поверхности разных предметов – на стволах деревьев, на заборах, на скалах, на снегу, на поверхности почвы и в ее толще.
В почве находят водоросли на глубине до 2,7 м, но большая часть их обитает в самых верхних слоях (до 1 см). Здесь они являются типичными фототрофами, однако в глубине почвы, в полной темноте, могут переходить на гетеротрофное питание.
В Мировом океане водоросли обитают в освещаемой зоне. Глубже всех проникают красные водоросли. Чаще они обитают на глубинах до 20–40 м, но если прозрачность воды велика, то встречаются до 100 и даже 200 м.
На суше для высших фотоавтотрофных растений условия освещения практически везде благоприятны, и они растут повсюду, где позволяют климатические и почвенные условия, приспосабливаясь к световому режиму данного местообитания.
3.2.2. Экологические группы растений по отношению к свету и их адаптивные особенности
Световой режим любого местообитания определяется интенсивностью прямого и рассеянного света, количеством света (годовой суммарной радиацией), его спектральным составом, а также альбедо – отражательной способностью поверхности, на которую падает свет.
Перечисленные элементы светового режима очень переменчивы и зависят от географического положения, высоты над уровнем моря, от рельефа, состояния атмосферы, характера земной поверхности, растительности, от времени суток, сезона года, солнечной активности и глобальных изменений в атмосфере.