2– отклонение оси «восьмерки» при передвижении солнца
Навигационная способность птиц врожденная. Она не приобретается за счет жизненного опыта, а создается естественным отбором как система инстинктов. Точные механизмы такой ориентации еще плохо изучены. Гипотеза ориентации птиц в перелетах по астрономическим источникам света в настоящее время подкреплена материалами опытов и наблюдений.
Способность к подобного рода ориентации свойственна и другим группам животных. Среди насекомых она особенно развита у пчел. Пчелы, нашедшие нектар, передают другим информацию о том, куда лететь за взятком, используя в качестве ориентира положение солнца. Пчела-разведчица, открывшая источник корма, возвращается в улей и начинает на сотах танец, совершая быстрые повороты. При этом она описывает фигуру в виде восьмерки, поперечная ось которой наклонена по отношению к вертикали. Угол наклона соответствует углу между направлениями на солнце и на источник корма (рис. 28). Когда медосбор очень обилен, разведчицы сильно возбуждены и могут танцевать долго, в течение многих часов, указывая сборщицам путь к нектару. За время их танца угол наклона восьмерки постепенно смещается в соответствии с движением солнца по небу, хотя пчелы в темном улье и не видят его. Если солнце скрывается за облаками, пчелы ориентируются на поляризованный свет свободного участка неба. Плоскость поляризации света зависит от положения солнца.
3.3. Влажность
Протекание всех биохимических процессов в клетках и нормальное функционирование организма в целом возможны только при достаточном обеспечении его водой – необходимым условием жизни. Поддержание водного баланса имеет огромное значение для всех живых организмов.
Проблемы водообеспечения особенно важны для обитателей суши. Особенности поддержания водного баланса зависят от того, в какой экологической обстановке они живут, какой образ жизни ведут, насколько могут использовать различные источники влаги и задерживать воду в теле.
3.3.1. Адаптация растений к поддержанию водного баланса
Низшие наземные растения из влажного субстрата поглощают воду погруженными в него частями таллома, а влагу дождя, росы и тумана – всей поверхностью. В максимально набухшем состоянии лишайники содержат в 20–30 раз больше воды, чем сухого вещества.
Среди высших наземных растений мохообразные поглощают воду из почвы ризоидами, а большинство других – корнями, специализированными органами, всасывающими воду. В клетках корня развивается сосущая сила чаще всего в несколько атмосфер, но этого достаточно для извлечения из почвы большей части связанной воды. Лесные деревья умеренной зоны развивают сосущую силу корней около 3 · 106 Па (30 атм), некоторые травянистые растения (земляника лесная, медуница неясная) – до 2 · 106 (20 атм) и даже свыше 4 · 106 Па (40 атм) (смолка обыкновенная); растения сухих областей – до 60 атм.
Когда в непосредственной близости от корней запасы воды в почве истощаются, корни растут в направлении большей влажности, так что корневая система растений постоянно находится в движении. У степных и пустынных растений часто можно видеть эфемерные корни, быстро вырастающие в периоды увлажнения почвы, а с наступлением засушливого периода засыхающие.
По типу ветвления различают следующие корневые системы:
1) экстенсивная охватывает большой объем почвы, но сравнительно слабо ветвится, так что почва пронизана корнями негусто. Таковы корневые системы у многих степных и пустынных растений (саксаула, верблюжьей колючки), у деревьев умеренной полосы (сосны обыкновенной, березы повислой), а из трав у люцерны серповидной, василька шероховатого и др.;
2) интенсивная– охватывает сравнительно небольшой объем почвы, но густо пронизывает ее многочисленными сильно ветвящимися корнями, как, например, у степных дерновинных злаков (ковылей, типчака и др.), у ржи, пшеницы. Между этими типами корневых систем есть переходные.
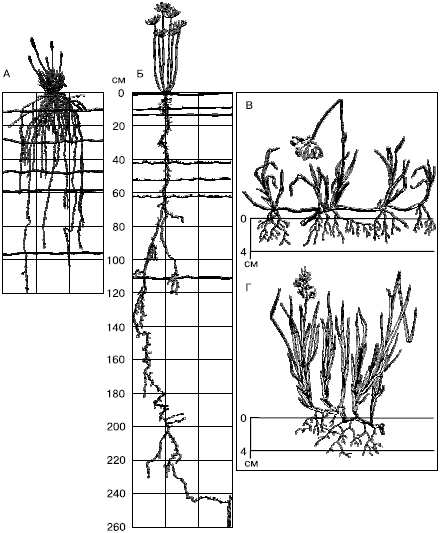
Корневые системы очень пластичны и резко реагируют на изменение условий, в первую очередь увлажнения (рис. 29). При недостатке влаги корневая система становится экстенсивнее. Так, при выращивании ржи в разных условиях общая длина корней (без корневых волосков) в 1000 см3 почвы варьирует от 90 м до 13 км, а поверхность корневых волосков может увеличиться в 400 раз.
Рис. 29. Корневые системы стенных и тундровых растений (по М. С. Шалыту и Б. А. Тихомирову, 1963):
А – Festuca sulcata;
Б – Euphorbia gerardiana на черноземах в Аскания-Нова;
В – Eriophorum scheuchzeri;
Г – Hierochloe alpina – из тундр Таймыра
Всасывание воды корнями затруднено при большой сухости почвы, засолении или сильной кислотности, при низкой температуре. Например, ясень обыкновенный при температуре почвы 0 °C поглощает воды в 3 раза меньше, чем при +(20–30) °С. Способность поглощать воду при той или иной температуре зависит от приспособленности растений к тепловому режиму почв в местах их произрастания. Виды с ранним началом развития, как правило, могут всасывать воду корнями при более низкой температуре, чем развивающиеся позднее. Тундровые растения и некоторые деревья, растущие на почвах с подстилающей их многолетней мерзлотой, могут поглощать воду при температуре почвы 0 °C.
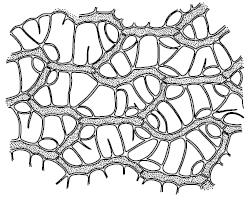
У высших растений есть и дополнительные пути поступления воды в тело. Мхи могут поглощать воду всей поверхностью, как и лишайники. Особенно много воды впитывают такие мхи, как кукушкин лен, виды сфагнума, чему способствует строение их листьев и побегов. При полном насыщении сфагновые мхи содержат в своем теле в десятки раз больше воды, чем в воздушно-сухом состоянии (рис. 30). Семена поглощают воду из почвы. Из воздуха, насыщенного водяными парами, в дождевом тропическом лесу поглощают воду многие эпифиты, например папоротник гименофиллум – тонкими листьями, многие орхидеи – воздушными корнями. В чашевидных влагалищах листьев многих зонтичных скапливается вода, которая постепенно всасывается эпидермисом. Виды из рода тилляндсия (бромелиевые) существуют в пустыне Атакама практически исключительно за счет влаги туманов и росы, которую впитывают чешуевидные волоски на листьях.
Поступившая в растение вода транспортируется от клетки к клетке (ближний транспорт) и по ксилеме во все органы, где расходуется на жизненные процессы (дальний транспорт). В среднем 0,5 % воды идет на фотосинтез, а остальная – на восполнение потерь от испарения и поддержание тургора. Вода испаряется со всех поверхностей, как внутренних, так и наружных, соприкасающихся с воздухом. Различают устьичную, кутикулярную и перидермальную транспирацию.
Рис. 30. Часть листа сфагнума болотного с клетками двух типов: зеленых с хлоропластами и бесцветных водоносных (Жизнь растений. Т. 4. 1978)
Через устьица транспирируется влага, испарившаяся с поверхности клеток внутри органов. Это основной путь расходования воды растением. Кутикулярная транспирация составляет менее 10 % от свободного испарения; у вечнозеленых хвойных пород она сокращается до 0,5 %, а у кактусов даже до 0,05 %. Относительно велика кутикулярная транспирация молодых развертывающихся листьев. Перидермальная транспирация обычно незначительна. Интенсивность общей транспирации повышается с увеличением освещенности, температуры, сухости воздуха и при ветре.
Водный баланс остается уравновешенным в том случае, если поглощение воды, ее проведение и расходование гармонично согласованы друг с другом. Нарушения его могут быть кратковременными или длительными. По приспособлениям наземных растений к кратковременным колебаниям условий водоснабжения и испарения различают пойкилогидрические и гомойогидрические виды.