Ослабление межвидовой конкуренции приводит к расширению экологической ниши вида. На океанических островах с бедной фауной ряд птиц по сравнению с их сородичами на материке заселяет более разнообразные местообитания и расширяет спектр кормов, так как не сталкивается при этом с конкурирующими видами. У островных обитателей отмечается даже повышенная изменчивость формы клюва как показатель расширения характера кормовых связей.
Если межвидовая конкуренция сужает экологическую нишу вида, не давая проявиться всем его потенциям, то внутривидовая конкуренция, наоборот, способствует расширению экологических ниш. При возросшей численности вида начинается использование дополнительных кормов, освоение новых местообитаний, появление новых биоценотических связей.
Экологические ниши растений, на первый взгляд, менее разнообразны, чем животных. Они четко очерчены у видов, различающихся по питанию: сапрофитов (подъельник, гнездовка), паразитов (заразиха, повилика, раффлезия), полупаразитов (погремок, марьянник, омела), насекомоядных растений (росянка, пузырчатка, непентес). Но и зеленые фотоавтотрофные растения занимают разные экологические ниши в фитоценозах. Множественность экологических ниш определяется разным использованием растениями среды, размещением органов в воздухе и почве, ритмами сезонного развития, длительностью периода вегетации, сроками цветения и особенностями плодоношения, взаимосвязями с элементами абиотической среды и всеми компонентами биоценоза.
В водоемах растения, полностью погруженные в воду (элодея, роголистник, уруть), оказываются в иных условиях температуры, освещенности, газового режима, чем плавающие на поверхности (телорез, водокрас, ряска) или укореняющиеся на дне и выносящие листья на поверхность (кувшинка, кубышка, виктория). Различаются они и взаимосвязями со средой. Эпифиты тропических лесов занимают сходные, но все же не идентичные ниши, так как относятся к разным экологическим группам по отношению к свету и воде (гелиофиты и сциофиты, гигрофиты, мезофиты и ксерофиты). Разные эпифитные орхидеи имеют узкоспециализированных опылителей.
В зрелом широколиственном лесу деревья первого яруса – дуб обыкновенный, вяз гладкий, клен платановидный, липа сердцелистная, ясень обыкновенный имеют сходные жизненные формы. Древесный полог, образованный их кронами, оказывается в одном горизонте, в сходных условиях среды. Но внимательный анализ показывает, что они по-разному участвуют в жизни сообщества и, следовательно, занимают разные экологические ниши. Эти деревья различаются по степени светолюбия и теневыносливости, срокам цветения и плодоношения, способам опыления и распространения плодов, составу консортов и проч. Дуб, вяз и ясень – анемофильные растения, но насыщение среды их пыльцой происходит в разные сроки. Клен и липа – энтомофилы, хорошие медоносы, но цветут в разное время. У дуба – зоохория, у остальных широколиственных деревьев – анемохория. Состав консортов у всех разный.
Если в широколиственном лесу кроны деревьев находятся в одном горизонте, то активные корневые окончания располагаются на разной глубине. Корни дуба проникают наиболее глубоко, выше располагаются корни клена и еще более поверхностно – ясеня. Опад разных видов деревьев утилизируется с разной скоростью. Листья липы, клена, вяза, ясеня к весне почти полностью разлагаются, а листья дуба и весной еще образуют рыхлую лесную подстилку.
В соответствии с представлениями Л. Г. Раменского об экологической индивидуальности видов и с учетом того, что виды растений в сообществе по-разному участвуют в освоении и преобразовании среды и трансформации энергии, можно считать, что в сложившихся фитоценозах каждый вид растения имеет свою экологическую нишу.
В онтогенезе растения, как и многие животные, меняют экологическую нишу. С возрастом они более интенсивно используют и преобразуют среду. Переход растения в генеративный период заметно расширяет круг консортов, меняет размер и напряженность фитогенного поля. Средообразующая роль стареющих, сенильных растений убывает. Они теряют многих консортов, но увеличивается роль связанных с ними деструкторов. Продукционные процессы ослабляются.
У растений имеет место перекрывание экологических ниш. Оно усиливается в отдельные периоды при ограничении ресурсов среды, но поскольку виды используют ресурсы индивидуально, избирательно и с разной интенсивностью, конкуренция в устойчивых фитоценозах ослабляется.
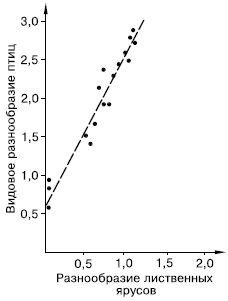
Рис. 94. Корреляция между разнообразием лиственных ярусов и видовым разнообразием птиц (индексы Шеннона по Макартуру из Э. Пианка, 1981)
На богатство экологических ниш в биоценозе оказывают влияние две группы причин. Первая – условия среды, предоставляемые биотопом. Чем мозаичнее и разнообразнее биотоп, тем больше видов могут размежевать в нем свои экологические ниши. Другой источник разнообразия ниш – сами виды, являющиеся ресурсом и создающие среду для других. Американский исследователь Р. Макартур, изучая ярусное разнообразие растительности по горному склону и число видов гнездящихся птиц, продемонстрировал прямую зависимость между этими показателями, положив начало многим исследованиям, подтверждающим связь видовой емкости сообществ с их внутренней структурой (рис. 94). Любой новый вид, внедряющийся в сообщество, увеличивает число имеющихся в нем экологических ниш не только за счет своего собственного положения среди других, но и предоставляя ресурсы для паразитов и хищников, норовых и гнездовых сожителей и т. п.
7.5. Ценотические стратегии видов
В фитоценологии разработаны классификации растений по их способности к совместному произрастанию и ценотической значимости. Общие положения этих классификаций могут быть применены и к животным, так как они характеризуют своего рода стратегии видов, определяющие их место в биоценозах. Наиболее часто используется система Л. Г. Раменского и Д. Грайма.
Группы растений, которые занимают сходное положение в фитоценозах, называют фитоценотипами. Л. Г. Раменский предложил различать среди совместно живущих растений три типа – виоленты, патиенты и эксплеренты. Он популярно характеризовал их как силовиков, выносливцев и выполняющих (т. е. заполняющих свободное пространство), уподобляя львам, верблюдам и шакалам. Виоленты обладают высокой конкурентной способностью в данных условиях: «энергично развиваясь, они захватывают территорию и удерживают ее за собой, подавляя, заглушая соперников энергией жизнедеятельности и полнотой использования ресурсов среды». Патиенты «в борьбе за существование… берут не энергией жизнедеятельности и роста, а своей выносливостью к крайне суровым условиям, постоянным или временным». Они довольствуются теми ресурсами, которые остаются от виолентов. Эксплеренты «имеют очень низкую конкурентную мощность, но зато они способны очень быстро захватывать освобождающиеся территории, выполняя промежутки между сильными растениями, так же легко они вытесняются последними».
Более подробные классификации выделяют и другие, промежуточные типы. В частности, можно различать еще группу пионерных видов, которые быстро занимают вновь возникающие территории, на которых еще не было никакой растительности. Пионерные виды частично обладают свойствами эксплерентов – низкой конкурентной способностью, но, как и патиенты, обладают высокой выносливостью к физическим условиям среды.
В 70-е годы прошлого века, спустя 40 лет после Л. Г. Раменского, выделение тех же трех фитоценотипов повторил незнакомый с его классификацией ботаник Д. Грайм, обозначив их другими терминами: конкуренты, толеранты и рудералы.
Практически в любой группе организмов выделяются аналогичные по способности к совместному существованию виды, поэтому классификацию ценотических стратегий Раменского-Грайма можно относить к общеэкологической.