Особый интерес представляют некоторые, сравнительно редкие виды, у которых способность к умерщвлению конкурентов внутри популяции закрепляется эволюционно в их поведении и даже морфологии. Подобные примеры встречаются среди насекомых.
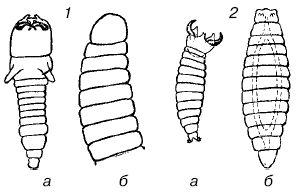
Для паразитоидных перепончатокрылых, откладывающих свои яйца в яйца или личинки других насекомых, запас пищи в хозяине весьма ограничен и дает возможность развиться чаще всего лишь одной особи паразита. У ряда видов личинки первого возраста имеют челюсти, ненужные при питании содержимым яйца, но приспособленные для схваток с конкурентами. Нападение личинок друг на друга обычно происходит до того, как они начинают кормиться. После уничтожения конкурента личинка линяет и ее острые челюсти сменяются мягкими и бугорковидными (рис. 130). У тех видов, самки которых способны распознать уже зараженных хозяев и не откладывать яйца в них повторно, личинкам не приходится конкурировать между собой, и они лишены острых челюстей.
Рис. 130. Личинки паразитоидных перепончатокрылых:
1– Opius fletcheri; 2 – Galesus sylvestrii (a – первый возраст, б – второй возраст)
Групповые паразиты, откладывающие по нескольку яиц в одно насекомое, нередко обладают способностью соразмерять число яиц с величиной жертвы. Однако при недостатке хозяев возможно перезаражение обнаруженных паразитами, при этом обеспеченность потомства кормом снижается. В таких случаях наблюдается уменьшение размеров личинок. Вышедшие из них имаго откладывают вдвое-втрое меньше яиц при сокращенных сроках жизни. Например, у самых мелких наездников-браконид происходит полная редукция яичников.
Плотность популяции у паразитических перепончатокрылых может регулироваться также через изменения в соотношении полов, что влияет на численность следующего поколения. У многих видов сильно различается смертность самцов и самок при перенаселении. Например, у некоторых браконид доля самок составляет около 70 % при слабом заражении насекомых-хозяев и падает ниже 10 % – при сильном. Кроме того, соотношение полов регулируется поведением самок. Для многих видов наездников установлено, что в мелких хозяевах, представляющих малый запас корма, самки откладывают преимущественно неоплодотворенные яйца, из которых у перепончатокрылых развиваются самцы. При учащении контактов самок друг с другом или при восприятии следовых запахов, оставляемых другими самками, среди откладываемых яиц также увеличивается доля неоплодотворенных.
Среди механизмов, задерживающих рост популяций, у многих видов большую роль играют химические взаимодействия особей. Так, вода аквариума, в котором содержались дафнии, способна задерживать рост представителей того же вида и сохраняет эту способность в течение нескольких дней. Головастики выделяют в воду частицы белковой природы, которые задерживают рост других головастиков. Чем крупнее особь, тем сильнее она воздействует на более мелких, так как устойчивость к одной и той же концентрации ингибитора находится в прямой зависимости от размеров. Один крупный головастик Rana pipiens может задержать рост всех других в 75-литровом аквариуме. Поколение, вышедшее в близкие сроки из икры, отложенной в одном водоеме, вскоре разделяется на две размерные группы: более крупные, продолжающие расти головастики и мелкие, затормозившие свой рост из-за неблагоприятной для них концентрации метаболита. Экологическая выгода такого разделения популяции в том, что особи с наследственно более быстрым темпом роста, используя в полной мере кормовые ресурсы водоема, получают возможность быстро завершить метаморфоз и в популяцию вливается полноценное пополнение. Оставшиеся мелкие головастики, после того как первая партия покинет водоем и в нем снизится концентрация ингибитора, также имеют шансы увеличиться в размерах и достичь стадии метаморфоза, но значительно позже. Эту вторую часть пополнения можно рассматривать как своего рода резерв, который вливается в популяцию лишь при достаточно благоприятных условиях (если не пересохнет временный водоем, если в нем продолжится воспроизводство водорослей – основной пищи головастиков и т. п.).
Выделение в окружающую среду продуктов, задерживающих рост, обнаружено у многих растений и водных животных, особенно у рыб.
Другой механизм ограничения численности популяций – такие изменения физиологии и поведения при увеличении плотности, которые в конечном счете приводят к проявлению инстинктов массовой миграции. В результате происходит выселение большей части популяции за пределы территории, занимаемой в оседлый период. Особенно ярко это проявляется у насекомых, которым свойственна фазовость – резкое изменение морфологии и физиологии особей в зависимости от плотности популяции (рис. 131). У перелетной саранчи-шистоцерки в постоянных очагах ее обитания в Индии, Пакистане, Восточной Африке и Аравии при низкой численности личинки одиночной фазы имеют ярко-зеленую, а взрослые – серовато-зеленую или бурую окраску. В годы массового размножения, которое наступает при благоприятном сочетании погодных условий, саранча переходит в стадную фазу. Личинки приобретают ярко-желтую, с черными пятнами окраску, взрослая неполовозрелая шистоцерка интенсивно розово-лиловая, половозрелая – лимонно-желтая. Изменяется и морфология особей: удлиняются надкрылья, меняются формы переднеспинки, киля, пропорции конечностей и т. п. Переход из одной фазы в другую занимает около трех поколений. Процесс стимулируется зрительным восприятием особой своего вида и контактами при помощи усиков. Это вызывает в организме насекомых целую серию реакций, приводящих к гормональным перестройкам, в которых участвуют эндокринные железы.
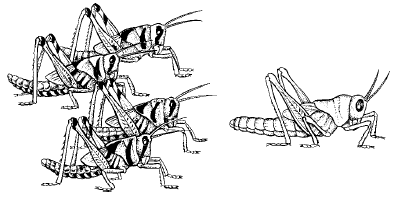
Рис. 131. Нимфы V возраста саранчи-шистоцерки (по Н. С. Щербиновскому, 1952): слева – стадная форма; справа – одиночная форма
Стадная фаза отличается повышенной возбудимостью и чрезвычайной прожорливостью. Плодовитость самок снижается, но они откладывают яйца с большим содержанием питательных веществ. Стадная саранча все время находится в состоянии миграционной активности. Личинки двигаются скоплениями – кулигами, а взрослые гигантскими стаями разлетаются на сотни и тысячи километров от мест постоянного обитания (рис. 132). Так, в конце прошлого столетия масса одной из стай шистоцерки, перелетевшей через Красное море, была определена не менее чем в 44 млн т.
На границах своего временного ареала перелетная саранча не может размножаться, и эти очаги вскоре затухают. Стаи или погибают, или, постепенно разреживаясь, начинают откочевку к зоне постоянных очагов. В изреженных популяциях вновь происходит переход к одиночной фазе, через 2–3 промежуточных поколения. Таким образом, разлет стай саранчи не обеспечивает формирования новых постоянных популяций, а практически служит лишь механизмом снятия перенаселенности в местах, благоприятных для размножения. При этом гибнет гигантское количество насекомых. Стаи, несущие на своем пути неисчислимые бедствия, сами оказываются обреченными.
Рис. 132. Нашествие перелетной саранчи
Явление фазовости обнаружено не только у стадных саранчовых, но и у других беспозвоночных. У тлей увеличение плотности населения вызывает появление крылатой фазы и разлет насекомых с образованием новых поселений. Обычно тли дают несколько поколений бескрылых самок, но в условиях постоянного перенаселения крылатые самки развиваются в каждом поколении. У ряда амеб химические изменения состава водной среды, вызванные переуплотнением популяции, стимулируют переход в подвижную жгутиковую стадию. В результате происходит быстрое рассредоточение особей в пространстве.