Формы конкурентного взаимодействия могут быть самыми различными: от прямой физической борьбы до мирного совместного существования. Тем не менее если два вида с одинаковыми экологическими потребностями оказываются в одном сообществе, рано или поздно один конкурент вытесняет другого. Это одно из наиболее общих экологических правил, которое получило название закона конкурентного исключения и было сформулировано Г. Ф. Гаузе.
В упрощенной форме оно звучит как «два конкурирующих вида вместе не уживаются».
Несовместимость конкурирующих видов еще раньше была подчеркнута Ч. Дарвином, который считал конкуренцию одной из важнейших составных частей борьбы за существование, играющей большую роль в эволюции видов.
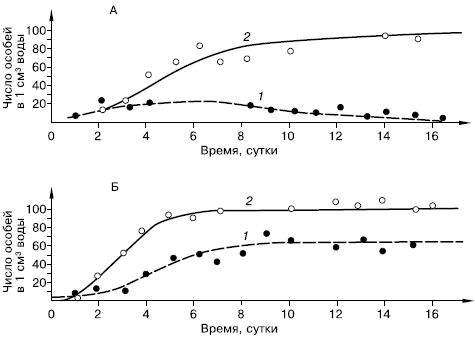
В опытах Г. Ф. Гаузе с культурами туфелек Paramecium aurelia и P. caudatum каждый из видов, помещенных отдельно в пробирки с сенным настоем, успешно размножался, достигая определенного уровня численности. Если же оба вида со сходным характером питания помещали совместно, то первое время наблюдался рост численности каждого из них, но затем количество P. caudatum постепенно сокращалось, и они исчезали из настоя, тогда как количество P. aurelia оставалось постоянным (рис. 86).
Рис. 86. Рост численности инфузорий Paramaecium caudatum (1) и P. aurelia (2) (по Г. Ф. Гаузе из Ф. Дре, 1976): А – в смешанной культуре; Б – в раздельных культурах
Победителем в конкурентной борьбе оказывается, как правило, тот вид, который в данной экологической обстановке имеет хотя бы небольшие преимущества перед другим, т. е. больше приспособлен к условиям окружающей среды, поскольку даже близкие виды никогда не совпадают по всему экологическому спектру. Так, в опытах Т. Паркас лабораторными культурами мучных хрущаков выявлено, что результат конкуренции может определяться тем, при какой температуре и влажности протекает опыт. В многочисленных стаканчиках с мукой, в которые помещали по нескольку экземпляров жуков двух видов (Tribolium confusum и Т. castaneum) и в которых они размножались, через некоторое время оставался только один из видов. При высокой температуре и влажности муки это был Т. castaneum, при более низкой температуре и умеренной влажности – Т. confusum. Однако при средних значениях факторов «победа» того или иного вида явно носила случайный характер, и предсказать исход конкуренции было трудно.
Причины вытеснения одного вида другим могут быть различны. Поскольку экологические спектры даже близких видов никогда не совпадают полностью, при общем сходстве требований к среде виды все же чем-либо отличаются друг от друга. Даже если такие виды мирно уживаются вместе, но интенсивность размножения одного чуть больше, чем другого, то постепенное исчезновение из сообщества второго вида лишь дело времени, так как с каждым поколением все больше и больше ресурсов оказывается захваченным более конкурентоспособным партнером. Часто, однако, конкуренты активно воздействуют друг на друга.
У растений подавление конкурентов происходит в результате перехвата минеральных питательных веществ и почвенной влаги корневой системой и солнечного света – листовым аппаратом, а также в результате выделения токсичных соединений. Например, в смешанных посевах двух видов клевера Trifolium repens раньше образует полог листьев, но затем его затеняет Т. fragiferum, у которого более длинные черешки. При совместном выращивании рясок Lemna gibba и Spirodela polyrrhiza численность второго вида сначала нарастает, а затем падает, хотя в чистых культурах скорость роста этого вида выше, чем первого. Преимущества L. gibba в данном случае в том, что в условиях загущения у него развивается аэренхима, помогающая держаться на поверхности воды. S. polyrrhiza, у которой нет аэренхимы, оттесняется вниз и затеняется конкурентом.
Химические взаимодействия растений через продукты их обмена веществ получили название аллелопатии. Подобные способы влияния друг на друга свойственны и животным. В приведенных выше опытах Г. Ф. Гаузе и Т. Парка подавление конкурентов происходило в основном в результате накопления в среде токсичных продуктов обмена, к которым один из видов более чувствителен, чем другой. Высшие растения с низкой потребностью в азоте, первыми появляющиеся на залежных почвах, корневыми выделениями подавляют образование клубеньков у бобовых и деятельность свободноживущих азотфиксирующих бактерий. Предотвращая тем самым обогащение почвы азотом, они получают преимущества в конкуренции с растениями, нуждающимися в большом его количестве в почве. Рогоз в зарастающих водоемах аллелопатически активен по отношению к другим водным растениям, что позволяет ему, избегая конкурентов, расти практически в чистых зарослях.
У животных могут встречаться случаи прямого нападения одного вида на другой в конкурентной борьбе. Например, личинки яйцеедов Diachasoma tryoni и Opius humilis, оказавшиеся в одном яйце хозяина, вступают друг с другом в схватку и убивают соперника, прежде чем приступить к питанию.
Возможность конкурентного вытеснения одного вида другим является результатом экологической индивидуальности видов. В неизменных условиях они будут иметь разную конкурентоспособность, так как обязательно отличаются друг от друга по толерантности к каким-либо факторам. В природе, однако, в большинстве случаев среда изменчива и в пространстве, и во времени, и это дает возможность сосуществования многих конкурентов. Например, если погодные условия более или менее регулярно меняются в пользу то одного, то другого вида, начинающиеся процессы вытеснения ими друг друга не доходят до конца и меняют знак на противоположный. Так, во влажные годы в нижнем ярусе леса могут разрастаться мхи, а в сухие их теснит покров осоки волосистой или других трав. Эти виды уживаются также в одном фитоценозе, занимая разные по условиям увлажнения участки леса. Кроме того, виды, конкурирующие не за один, а за несколько ресурсов, часто имеют разные пороги ограничивающих факторов, что также мешает завершиться процессам конкурентного исключения. Так, американский эколог Д. Тилман, культивируя совместно два вида диатомовых водорослей, выяснил, что они не вытесняют друг друга, потому что имеют разную чувствительность к недостатку азота и кремния. Вид, способный в размножении опередить другой при низком содержании азота, не может этого достичь из-за нехватки для него кремния, тогда как его конкуренту, наоборот, достаточно кремния, но мало азота.
Конкурирующие виды могут уживаться в сообществе и в том случае, если повышение численности более сильного конкурента не допускается хищником. В этом случае деятельность хищника приводит к повышению видового разнообразия сообщества. В одном из опытов со дна прибрежного участка моря, где обитало 8 видов сидячих беспозвоночных – мидий, морских желудей, морских уточек, хитонов, – удалили хищника, морскую звезду, питавшуюся в основном мидиями. Через некоторое время мидии заняли весь участок дна, вытеснив все остальные виды.
Таким образом, биоценозы содержат в каждой группе организмов значительное число потенциальных или частичных конкурентов, состоящих в динамических отношениях друг с другом. Вид может не иметь также сильных соперников, но испытывать небольшое влияние со стороны каждого из многих других, частично использующих его ресурсы. В этом случае говорят о «диффузной» конкуренции, исход которой также зависит от многих обстоятельств и может закончиться вытеснением данного вида из биоценоза.
Конкуренция, следовательно, имеет двоякое значение в биоценозах. Она является фактором, в значительной мере определяющим видовой состав сообществ, поскольку интенсивно конкурирующие виды вместе не уживаются. С другой стороны, частичная или потенциальная конкуренция позволяет видам быстро захватывать дополнительные ресурсы, освобождающиеся при ослаблении деятельности соседей, и замещать их в биоценотических связях, что сохраняет и стабилизирует биоценоз в целом.