Если вода гипертонична по отношению к жидкостям тела гидробионтов, им грозит обезвоживание в результате осмотических потерь. Защита от обезвоживания достигается повышением концентрации солей также в теле гидробионтов. Обезвоживанию препятствуют непроницаемые для воды покровы гомойосмотических организмов – млекопитающих, рыб, высших раков, водных насекомых и их личинок.
Многие пойкилосмотические виды переходят к неактивному состоянию – анабиозу в результате дефицита воды в теле при возрастании солености. Это свойственно видам, обитающим в лужах морской воды и на литорали: коловраткам, жгутиковым, инфузориям, некоторым рачкам, черноморским полихетам Nereis divesicolor и др. Солевой анабиоз– средство переживать неблагоприятные периоды в условиях переменной солености воды.
Истинно эвригалинных видов, способных в активном состоянии обитать как в пресной, так и в соленой воде, среди водных обитателей не так много. В основном это виды, населяющие эстуарии рек, лиманы и другие солоноватоводные водоемы.
Температурный режим водоемов более устойчив, чем на суше. Это связано с физическими свойствами воды, прежде всего высокой удельной теплоемкостью, благодаря которой получение или отдача значительного количества тепла не вызывает слишком резких изменений температуры. Испарение воды с поверхности водоемов, при котором затрачивается около 2263,8 Дж/г, препятствует перегреванию нижних слоев, а образование льда, при котором выделяется теплота плавления (333,48 Дж/г), замедляет их охлаждение.
Амплитуда годовых колебаний температуры в верхних слоях океана не более 10–15 °C, в континентальных водоемах – 30–35 °C. Глубокие слои воды отличаются постоянством температуры. В экваториальных водах среднегодовая температура поверхностных слоев +(26–27) °С, в полярных – около 0 °C и ниже. В горячих наземных источниках температура воды может приближаться к +100 °C, а в подводных гейзерах при высоком давлении на дне океана зарегистрирована температура +380 °C.
Таким образом, в водоемах существует довольно значительное разнообразие температурных условий. Между верхними слоями воды с выраженными в них сезонными колебаниями температуры и нижними, где тепловой режим постоянен, существует зона температурного скачка, или термоклина. Термоклин резче выражен в теплых морях, где сильнее перепад температуры наружных и глубинных вод.
В связи с более устойчивым температурным режимом воды среди гидробионтов в значительно большей мере, чем среди населения суши, распространена стенотермность. Эвритермные виды встречаются в основном в мелких континентальных водоемах и на литорали морей высоких и умеренных широт, где значительны суточные и сезонные колебания температуры.
Световой режим. Света в воде гораздо меньше, чем в воздухе. Часть падающих на поверхность водоема лучей отражается в воздушную среду. Отражение тем сильнее, чем ниже положение Солнца, поэтому день под водой короче, чем на суше. Например, летний день около острова Мадейра на глубине 30 м – 5 ч, а на глубине 40 м всего 15 мин. Быстрое убывание количества света с глубиной связано с поглощением его водой. Лучи с разной длиной волны поглощаются неодинаково: красные исчезают уже недалеко от поверхности, тогда как сине-зеленые проникают значительно глубже. Сгущающиеся с глубиной сумерки в океане имеют сначала зеленый, затем голубой, синий и сине-фиолетовый цвет, сменяясь наконец постоянным мраком. Соответственно сменяют друг друга с глубиной зеленые, бурые и красные водоросли, специализированные на улавливании света с разной длиной волны.
Окраска животных меняется с глубиной так же закономерно. Наиболее ярко и разнообразно окрашены обитатели литоральной и сублиторальной зон. Многие глубинные организмы, подобно пещерным, не имеют пигментов. В сумеречной зоне широко распространена красная окраска, которая является дополнительной к сине-фиолетовому свету на этих глубинах. Дополнительные по цвету лучи наиболее полно поглощаются телом. Это позволяет животным скрываться от врагов, так как их красный цвет в сине-фиолетовых лучах зрительно воспринимается как черный. Красная окраска характерна для таких животных сумеречной зоны, как морской окунь, красный коралл, различные ракообразные и др.
У некоторых видов, обитающих у поверхности водоемов, глаза разделяются на две части с разной способностью к преломлению лучей. Одна половина глаза видит в воздухе, другая – в воде. Такая «четырехглазость» характерна для жуков-вертячек, американской рыбки Anableps tetraphthalmus, одного из тропических видов морских собачек Dialommus fuscus. Эта рыбка при отливах сидит в углублениях, выставляя часть головы из воды (см. рис. 26).
Поглощение света тем сильнее, чем меньше прозрачность воды, которая зависит от количества взвешенных в ней частиц.
Прозрачность характеризуют предельной глубиной, на которой еще виден специально опускаемый белый диск диаметром около 20 см (диск Секки). Самые прозрачные воды – в Саргассовом море: диск виден до глубины 66,5 м. В Тихом океане диск Секки виден до 59 м, в Индийском – до 50, в мелких морях – до 5-15 м. Прозрачность рек в среднем 1–1,5 м, а в самых мутных реках, например в среднеазиатских Амударье и Сырдарье, всего несколько сантиметров. Граница зоны фотосинтеза поэтому сильно варьирует в разных водоемах. В самых чистых водах эуфотическая зона, или зона фотосинтеза, простирается до глубин не свыше 200 м, сумеречная, или дисфотическая, зона занимает глубины до 1000–1500 м, а глубже, в афотическую зону, солнечный свет не проникает совсем.
Количество света в верхних слоях водоемов сильно меняется в зависимости от широты местности и от времени года. Длинные полярные ночи сильно ограничивают время, пригодное для фотосинтеза, в арктических и приантарктических бассейнах, а ледовый покров затрудняет доступ света зимой во все замерзающие водоемы.
В темных глубинах океана в качестве источника зрительной информации организмы используют свет, испускаемый живыми существами. Свечение живого организма получило название биолюминесценции. Светящиеся виды есть почти во всех классах водных животных от простейших до рыб, а также среди бактерий, низших растений и грибов. Биолюминесценция, по-видимому, многократно возникала в разных группах на разных этапах эволюции.
Химия биолюминесценции сейчас довольно хорошо изучена. Реакции, используемые для генерации света, разнообразны. Но во всех случаях это окисление сложных органических соединений (люциферинов) с помощью белковых катализаторов (люцифераз). Люциферины и люциферазы у разных организмов имеют неодинаковую структуру. В ходе реакции избыточная энергия возбужденной молекулы люциферина выделяется в виде квантов света. Живые организмы испускают свет импульсами, обычно в ответ на раздражения, поступающие из внешней среды.
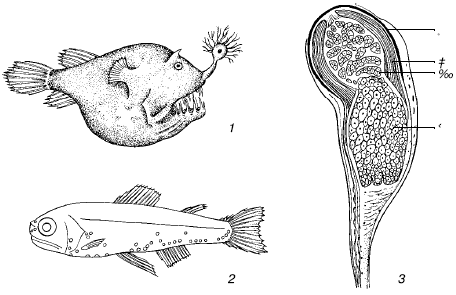
Свечение может и не играть особой экологической роли в жизни вида, а быть побочным результатом жизнедеятельности клеток, как, например, у бактерий или низших растений. Экологическую значимость оно получает только у животных, обладающих достаточно развитой нервной системой и органами зрения. У многих видов органы свечения приобретают очень сложное строение с системой отражателей и линз, усиливающих излучение (рис. 40). Ряд рыб и головоногих моллюсков, неспособных генерировать свет, используют симбиотических бактерий, размножающихся в специальных органах этих животных.
Рис. 40. Органы свечения водных животных (по С. А. Зернову, 1949):
1– глубоководный удильщик с фонариком над зубатой пастью;
2– распределение светящихся органов у рыбы сем. Mystophidae;
3– светящийся орган рыбы Argyropelecus affinis:
а – пигмент, б – рефлектор, в – светящееся тело, г – линза
Биолюминесценция имеет в жизни животных в основном сигнальное значение. Световые сигналы могут служить для ориентации в стае, привлечения особей другого пола, подманивания жертв, для маскировки или отвлечения. Вспышка света может быть защитой от хищника, ослепляя или дезориентируя его. Например, глубоководные каракатицы, спасаясь от врага, выпускают облако светящегося секрета, тогда как виды, обитающие в освещенных водах, используют для этой цели темную жидкость. У некоторых донных червей – полихет – светящиеся органы развиваются к периоду созревания половых продуктов, причем светятся ярче самки, а глаза лучше развиты у самцов. У хищных глубоководных рыб из отряда удильщиковидных первый луч спинного плавника сдвинут к верхней челюсти и превращен в гибкое «удилище», несущее на конце червеобразную «приманку» – железу, заполненную слизью со светящимися бактериями. Регулируя приток крови к железе и, следовательно, снабжение бактерии кислородом, рыба может произвольно вызывать свечение «приманки», имитируя движения червя и подманивая добычу.