В прошлом специалисты выделяли два основных вида диких предков крупного рогатого скота: дикий евро-африканский тур (Bos primigenius) и найденный лишь в плейстоценовых отложениях Индии азиатский (индийский) бык (Bos nomadicus) [7, с. 8—23]. В настоящее время считается, что у крупного рогатого скота был один дикий видовой предок В. primigenius, который включал ряд подвидов: европейский В.р. primigenius Bojanus, азиатский В.р. nomadicus Falconer et Cautley, североафриканский B.p.opisthonomus Pomel и камбоджийский B.p.sauveli [1056, c. 160; 1057, c. 201; 563, I, c. 226; 351, c. 126]. По сути дела, это чисто географическое разделение, так как внутри подвидов наблюдается значительное разнообразие [1057, с. 207; 563, I, с. 226]. Считается, что обыкновенный крупный рогатый скот произошел от европейского и североафриканского подвидов, однако не все детали этого процесса достаточно ясны, вызывая споры, касающиеся в особенности роли африканской разновидности [ср. 7, с. 12, 13; 429, с. 106; 563, I, с. 235–241]. От В.р. nomadicus происходит индийский горбатый скот, зебу.
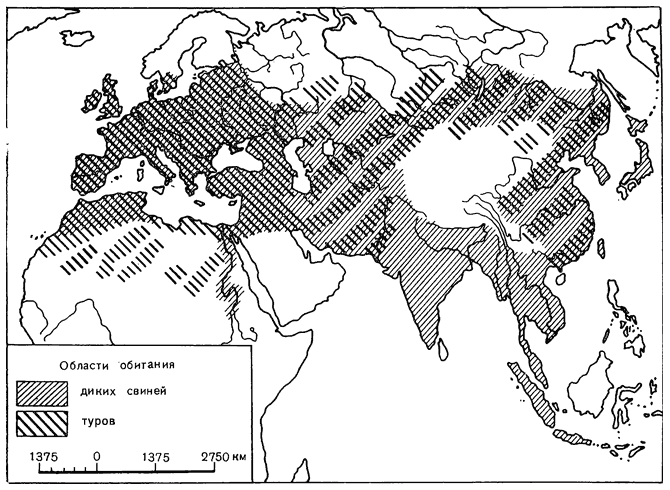
Области обитания кабанов и туров:
1 — ареал кабанов; 2 — ареал туров (прерывистой линией обозначены малоизученные районы)
Тур теперь полностью вымер. В древности он обитал от Тихого до Атлантического океана, от северной тундры до Индии и Северной Африки. Трудно что-либо точно сказать о его повадках и особенностях обитания. По всей видимости, тур предпочитал увлажненные участки степей и открытых лиственных лесов с умеренным и теплым климатом. Вместе с тем его широкое распространение говорит о способности приспосабливаться к весьма различной природной обстановке. Так, родоначальником зебу стал пустынный подвид тура, хорошо переносивший сухой жаркий климат и живший, по мнению Г. Эпштейна, на окраинах (пустынь Гельманд, Лут и Большой Солончаковой [563, I, с. 518]. Летом туры питались травой, молодыми побегами и листьями, осенью большую роль в их, питании играли желуди, а зимой им приходилось довольствоваться сухими ветвями и листвой. Тур представлял собой сильного, быстрого и свирепого зверя, и приручить его было довольно сложно [303, с. 600–616].
Точная систематика диких свиней до сих пор не выработана. В прошлом наблюдалась тенденция выделять несколько видов этих животных. Л. Адамец называл три вида: северо- и центральноевропейский (Sus scrofa ferns), средиземноморский (S. mediterraneus) и азиатский вид Южной и Восточной Азии (S. vittatus), [7, с. 45–48]. Г. П. Адлерберг объединял европейских, средиземноморских и восточноазиатских свиней в один вид (Sus scrofa), выделяя внутри него ряд подвидов. Он считал, что дикие свиньи Южной Азии относятся к особым видам: S.cristatus и S.vittatus [8, с. 202–206]. Подобным же образом Ф. Цейнер еще недавно выделял в особый вид Sus scrofa с подвидами S.s. scrofa (кабаны Северной Европы), S.s. meridionalis (средиземноморский подвид, включающий также и североафриканского S.s. barbarus) и S.s. attila (евро-азиатские кабаны), а также вслед за Г. П. Адлербергом он группировал свиней Юго-Восточной Азии в два особых вида. По мнению Ф. Цейнера, европейские домашние свиньи произошли от S. scrofa, а китайские от S.vittatus [1057, с. 256]. Сейчас считается, что все современные дикие свиньи относятся к одному виду S. scrofa, который делится на ряд подвидов [897, с. 139; 563, II, с. 316]. Так, Г. Эпштейн выделяет следующие подвиды: свиньи Юго-Восточной Азии (S.s.vittatus и S.s. cristatus), Центральной Европы (S.s.scrofa), Северной Африки (S.s.barbarus) и Северо-Восточной Африки (S.s. sennaarensis) [563, II, 316]. Впрочем, общепринятой внутривидовой систематики до сих пор нет.
Дикие свиньи ныне широко распространены от Атлантического океана до Тихого и обитают в самых разнообразных природных условиях. Они населяют долины и дельты больших и малых рек, приморские низменности, леса, горы вплоть до альпийской зоны. В определенные сезоны они не избегают и пустынных ландшафтов. Все же, где бы они ни обитали, они всегда держатся вблизи сырых болотистых мест, у водоемов. Для укрытия свиньи используют кустарники, заросли камыша, молодую поросль хвойного леса. В горах они придерживаются лесной зоны. В случае опасности животные мчатся напролом и свободно проходят там, где не только человеку, но и собаке пройти невозможно. В горных районах жизнь свиней отличается сезонностью. Лето они проводят в нижнем поясе леса, зимой часть популяции (молодые и холостые самцы) поднимается высоко в горы. В конце лета — осенью кабаны концентрируются в рощах диких фруктовых и ореховоплодных деревьев. Кабаны — всеядные животные, однако они предпочитают травянистые растения (злаки и др.), которые составляют их основную пищу весной — в начале лета. Дикие свиньи держатся, как правило, небольшими группами, редко превышающими 10–20 голов [303, с. 74–96; 637, с. 372–376]. Свиньи Южной и Восточной Азии отличаются скороспелостью и способностью к откорму, а свиньи Европы — крупными размерами и плодовитостью [7, с. 47]. Пойманные в молодом возрасте поросята диких свиней легко становятся ручными [303, с. 96].
Так как одним из важных источников питания всех названных животных являются дикие злаки, неудивительно, что ареалы тех и других во многом совпадают. Для диких хлебных злаков границы ареалов в последние годы были установлены довольно точно [1060, с. 48–66]. Дикая однозернянка (Triticum boeoticum) в своих двух разновидностях встречается от Балкан до Палестины и гор Загроса, эммер (Tr. dicoccoides) концентрируется в Леванте, а ячмень (Hordeum spontaneum) растет от Палестины до Юго-Западного Ирана. Все эти растения тяготеют к дубово-фисташковой лесостепи, однако местами заходят глубоко в степи, что особенно характерно для засухоустойчивого ячменя. В этих же районах локализуется первичный ареал диких бобовых, которые наряду со злаками составили тот биологический фонд, который лег в основу древнейшего земледелия [1061, с. 887–893]. Совпадение областей обитания диких сородичей домашних животных и культурных растений во многом объясняет то, почему производящее хозяйство возникло в Передней Азии как комплексное, сочетающее в себе земледелие и скотоводство.
Данные о современном расположении растительных зон имеют лишь относительную ценность для изучения процесса перехода к производящему хозяйству, так как начиная с конца плейстоцена их границы колебались как в связи с климатическими изменениями, так и в связи с деятельностью человека [691, с. 21–25]. Поэтому огромное значение имеет вопрос о реконструкции природной обстановки конца плейстоцена — начала голоцена. К сожалению, данных для изучения древнего климата в Передней Азии еще немного, и они зачастую трактуются по-разному. Все же сопоставление результатов, полученных разными методами независимо друг от друга (изучение морских и озерных отложений, атмосферных данных, пыльцевых диаграмм, микрофауны и т. д.), позволяет представить общую картину изменений природных условий в конце плейстоцена и в раннем и среднем голоцене (обзорные работы см. [372; 1049; 570; 468, с. 547–551; 513; 1052; 840]).
Теперь можно считать окончательно установленным, что примерно с XIV (XIII) тысячелетия до н. э. наблюдался длительный период потепления. Если в начале этого периода среднегодовая температура в Передней Азии была на 5–7° ниже современной [570, с. 557; 840, с. 12], то 10 (9) тыс. лет назад она достигла современного уровня, а 6,5 (6) тыс. лет назад она превысила его на 3° [513, с. 320; 840, с. 19]. Аналогичная последовательность отмечается и при изучении уровня влажности с той лишь разницей, что изменения влажности следовали за температурными изменениями с некоторым отставанием (в несколько сотен или тысяч лет?) [840, с. 15, 16]. Процесс потепления не был непрерывным. Временами он сменялся относительно короткими периодами похолодания, которые, правда, не были столь резко выражены, как в Европе, и поэтому фиксируются не во всех районах Передней Азии. Лучше всего сейчас известно похолодание, наступившее в позднем дриасе в X (IX) тысячелетии до н. э. В это время в Анатолии наблюдалось короткое наступление ледника [798, с. 222], а в Палестине и Южном Прикаспии проходила очередная регрессия [786, с. 391–399; 983, с. 40]. Пыльцевая диаграмма из пещеры Шанидар тоже свидетельствует о более холодном и влажном климате в этот период [947, с. 736]. Изучение микрофауны — с некоторых памятников Палестины, а также пыльцевой анализ, проведенный в пустыне Негев, показывают, что по крайней мере частично натуфийцы жили в период, более влажный, чем сейчас [983, с. 36, 39]. Наконец, данные, собранные У Нютцелем, также подтверждают, что в X (IX) тысячелетии до н. э. в Передней Азии наблюдалось временное похолодание, хотя количество осадков продолжало расти [840, с. 13–17].