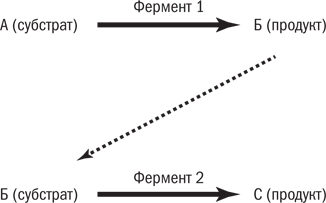
Относительные размеры ферментов очень велики. Их молекулы могут быть в 10–20 тыс. раз больше молекул субстрата, который они обрабатывают. И правда похоже на фабрику и полено. На рис. 7.4 показан субстрат А, превращающийся в продукт Б. Однако большинство реакций не происходит изолированно: они сопряжены с последующими, где Б (теперь уже субстрат) превращается в В (новый продукт). Фермент 1 превращает А в Б, а фермент 2 – Б в В.
Рис. 7.4. Простая ферментативная реакция
Ферменты могут работать с разной силой в зависимости от запасов (количества субстрата) и потребностей (количества имеющегося в клетке продукта). Как конвейер, который движется быстрее или медленнее в зависимости от поставки сырья и спроса на готовую продукцию, ферменты меняют скорость превращения субстратов (на профессиональном языке – «активность»). Они могут катализировать даже обратные реакции, превращая продукт в субстрат. В общем, от ферментов зависит, произойдет ли реакция, а если да, то как быстро и в каком направлении.
Исходная форма ферментов напоминает цепочку аминокислот, расположенных в последовательности, которая закодирована в ДНК. Но, поскольку аминокислоты имеют химическое и физическое сродство, цепочка складывается и образует трехмерную форму, как очень длинная нить намагниченных бусин (рис. 7.5).
Рис. 7.5. Компьютерная модель фермента цАДФ-рибозы-гидролазы (CD38)
Один из способов корректировки ферментативной активности – изменение формы фермента. Это имеет серьезные последствия, потому что меняет его химические и физические свойства, а также способность модифицировать скорость реакции. Многие ученые-энзимологи поэтизируют быстроту, с которой ферменты меняют конфигурацию для выполнения своих задач. Вот показательная статья из New World Encyclopedia (http://www.newworldencyclopedia.org):
Чтобы фермент был функционален, он должен принять трехмерную форму. Как происходит этот сложный процесс, остается загадкой. Небольшая цепочка из 150 аминокислот образует фермент, имеющий невероятное число возможных конфигураций: если проверять по 1012 разных конфигураций в секунду, потребуется 1026 лет, чтобы найти верную… Но денатурировавший фермент может правильно сложиться за долю секунды, а затем участвовать в химических реакциях… [Это] показывает ошеломляющую сложность и гармонию Вселенной{58}.
Пытаясь описать неописуемое, автор приводит пример сравнительно небольшой (для фермента) гипотетической молекулы. Скорость складывания фермента из линейной цепочки в готовую к работе сферу феноменальна. Не менее потрясает химическое разнообразие субстратов, которые может метаболизировать один активный фермент. И так же впечатляет огромное число факторов, способных модифицировать структуру ферментов, их число и активность.
Все это показывает глубокую связь между метаболизмом питательных веществ и миром ферментов. Катализируемые ими реакции, бесконечные числом и бесконечно переплетенные, контролируются нутриентами и связанными соединениями, число которых тоже бесконечно. Питательные вещества контролируют ферменты, и последние также действуют на питательные вещества, образуя нескончаемые продукты, которые затем своевременно используются для правильной работы организма.
Парадокс ОСФ
И мы наконец возвращаемся к ОСФ и его роли в образовании рака.
Здесь я вынужден подытожить, сократить и упростить наши исследования и открытия: тема слишком обширна и специфична, чтобы объяснить ее в одной главе. Моя цель – не сделать вас экспертом по ОСФ. Рассказывая о своем более чем пятидесятилетнем научном приключении с этим ферментом, я надеюсь, что вы лучше поймете, как животный белок влияет на образование рака, и глубже осознаете, что сложность ОСФ красноречиво подтверждает холистическое, а не редукционистское видение питания и здоровья.
ОСФ – исключительно сложный фермент, метаболизирующий многие субстраты, одни из которых обычно присутствуют в организме, а с другими он сталкивается впервые. ОСФ расположен в основном (но не только) в печени и метаболизирует стероидные гормоны (например, половые – эстрогены, андрогены и стрессовые), жирные кислоты (например, прекурсоры веществ, поддерживающих иммунную и нервную системы) и холестерин (вызывающий сердечно-сосудистые заболевания и являющийся частью клеточных мембран), а также другие соединения, образуя вещества, более близкие к тем, которые использует наш организм. ОСФ также обезвреживает инородные химические вещества, благодаря чему они легче выводятся из организма с мочой.
В начале моей научной карьеры меня учили, что афлатоксин (как и другие канцерогены) преобразуется ОСФ в менее токсичный метаболит, который выводится с мочой и калом. Это происходит следующим образом (рис. 7.6).
Рис. 7.6. Предполагаемая модель преобразования афлатоксина ОСФ
Однако модель явно упрощена. Во-первых, упомянутые индийские исследователи, опубликовавшие в 1968 году свои изыскания о высокобелковой (20 % белка) диете, усиливающей афлатоксин-индуцированные опухоли у крыс{59}, показали, что та же диета уменьшает вред афлатоксина, если давать его в очень высоких дозах{60}. Это был парадокс, который традиционная модель метаболизма афлатоксина объяснить не могла.
Подозревая, что ключ к решению – ОСФ, мы с коллегами установили, что высокобелковая диета повышает активность данного фермента у крыс{61}. Это значило, что чем больше белка ели крысы, тем быстрее происходило обезвреживание афлатоксина (точнее, его исходного субстрата – AFB1). Это открытие имело смысл, но не согласовывалось с наблюдением индийских ученых{62}, что высокобелковая диета повышает заболеваемость раком.
Один из рассматриваемых нами вариантов – ОСФ может образовывать два вида метаболитов: один – менее токсичный, чем афлатоксин, и безопасно выводимый из организма, другой – более токсичный и ведущий к раку. Но почему фермент действует так противоречиво? На первый взгляд странно, но вполне возможно. Задолго до этого и до открытия ОСФ ученые полагали, что многие химические канцерогены вызывают рак только после «активации» ферментами, и то, что вещества вроде афлатоксина дают более токсичный метаболит, звучало вполне правдоподобно.
Другой ключ к загадке был найден в начале 1970-х, когда профессора Висконсинского университета, видные онкологи Джим и Бетти Миллер вместе с коллегой Колином Гарнером, получили интересные данные: образование ОСФ нетоксичного метаболита афлатоксина проходит с образованием исключительно реактивного канцерогенного промежуточного продукта{63}. Иными словами, из афлатоксина ОСФ вырабатывает два метаболита: один обезвреживается и выводится из организма, а другой активируется и вызывает рак. Это как если подать на фабрику полено и получить на долю секунды полицейскую дубинку, которая позже примет форму салатницы.
Этот промежуточный метаболит известен как эпоксид. Считается, что он существует всего несколько миллисекунд, но этого, к сожалению, достаточно, чтобы он успел связаться с клеточным ДНК и привести к мутации, способной вызвать каскад событий, ведущих к раку.