В 1983 г. Освальд Стюарт и Уильям Леви исследовали[1632] влияние на синаптическую пластичность относительной синхронизации в пределах нескольких миллисекунд потенциалов пре- и постсинаптического действия. Для этого они взяли два нервных пути, оканчивающихся в одном постсинаптическом нейроне, — «слабый» и «сильный», то есть с меньшей эффективностью и с большей, — и вызывали в них стимулы, варьируя время вызова импульса. Хотя им и удалось обнаружить в опытах существенную асимметрию: активация слабого нервного пути раньше сильного вызывала ДВП в слабом нервном пути, в то время как активация сильного нервного пути раньше слабого вызывала ДВД в слабом нервном пути, но авторы, как и многие другие исследователи в 1980-е гг., не усмотрели в этом явлении системы.
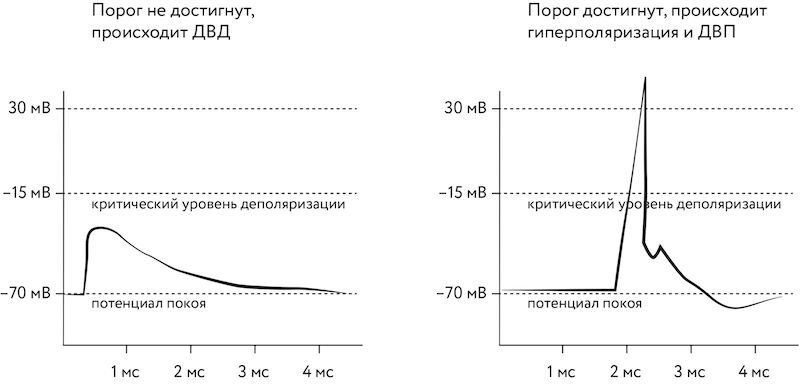
В 1990 г. немецкий исследователь Вольф Зингер и его коллеги сообщили[1633], что наступление ДВП или ДВД обусловлено гиперполяризацией или деполяризацией постсинаптического нейрона (напомним, что гиперполяризация — это смещение мембранного потенциала относительно потенциала покоя в более отрицательную сторону, а деполяризация — в положительную). ДВП происходит, если после стимуляции мембранный потенциал постсинаптического нейрона не достигает определённого порога («критический уровень деполяризации»), а ДВП происходит, если данный порог достигается, ввиду чего происходит последующая гиперполяризация. Рис. 115. Зависимость мембранного потенциала от времени, прошедшего после стимуляции Это исследование привлекло внимание к более пристальному изучению постсинаптического нейрона, поскольку именно его мембранный потенциал является ключевым для ДВП или ДВД, которые необязательно зависят от поступления сигнала через входящий синапс[1634]. Четырьмя годами спустя Доминик Дебанн и его коллеги сделали[1635] ещё один шаг вперёд: они вначале направляли деполяризующие импульсы тока непосредственно в постсинаптический нейрон, меняя его мембранный потенциал, а потом стимулировали нервный путь. Их исследования показали, что при небольшой разнице по времени между искусственной деполяризацией и входными импульсами происходит ДВП, а при большой — ДВД. Это добавило к исследованию Зингера тот факт, что именно уровень деполяризации и гиперполяризации, вызванный каким-либо образом (даже искусственно), и определяет направление синаптической пластичности. Наконец, в 1991 г. Роберто Малиноу провёл завершающее исследование. Он смог в тонком срезе гиппокампа выделить четыре пары пирамидальных нейронов CA3 и CA1, каждая из которых была связана через единственный синапс. Затем он, одновременно вызывая серии импульсов в пре- и постсинаптическом нейронах, получил ДВП в этих соединениях[1636]. Эта работа стала первой (спустя более сорока лет после выхода в свет ключевой работы Хебба!) прямой демонстрацией того, что синаптические связи между двумя нейронами могут изменяться, ведь здесь удалось полностью исключить влияние на ход эксперимента сторонних воздействий. В наши дни продолжается активное изучение биологических механизмов, лежащих в основе ДВП и ДВД. В частности, учёные вырабатывают подходы, позволяющие рассчитывать эффективность каждого отдельно взятого синапса на основе данных микроскопии[1637], [1638], [1639]. Важной вехой в истории изучения механизмов ДВП и ДВД стало выступление Генри Маркрама на ежегодном собрании Общества нейронаук (Society for Neuroscience) в 1995 г. В ходе этого выступления Маркрам рассказал научной общественности о первом экспериментальном исследовании[1640], показавшем, что ДВП и ДВД зависят от разницы во времени возникновения одиночных импульсов, испускаемых пре- и постсинаптическими нейронами в моносинаптических связях между парами нейронов неокортекса. Водораздел, задаваемый относительной синхронизацией отдельных импульсов на временно́м масштабе в несколько десятков миллисекунд, определял направление (потенциация или депрессия) и степень изменения эффективности синаптической связи. Именно это выступление Маркрама (ныне директора Blue Brain Project и основателя Human Brain Project) стало моментом окончательного оформления STDP в качестве модели синаптической пластичности биологических нейронов и одновременно как алгоритма обучения импульсных нейронных сетей[1641], хотя сам термин STDP появился только в 2000 г. в работе[1642] Сена Сонга, Кеннета Миллера и Ларри Эбботта. Таким образом, спустя полстолетия после основополагающей работы Хебба его знаменитое «возбуждаются вместе — связываются вместе» [fire together, wire together] (соответствующее ДВП), как оказалось, можно дополнить ещё одним принципиальным афоризмом, предложенным Карлой Шатц: «Сработал несинхронно — потерял свою связь» [fire out of sync, lose your link][1643] (соответствующее ДВД). Важными параметрами STDP являются конкретные значения временных интервалов синхронизации импульсов. В ходе экспериментов по обучению больших импульсных сетей для решения прикладных задач были созданы такие методы, как R-STDP (версия STDP, в которой обучение модулируется при помощи внешнего подкрепления, основанного на фазовой активности дофаминовых нейронов)[1644], TD-STDP (версия STDP, использующая метод временных разниц [temporal difference])[1645] и даже BP-STDP, а также другие похожие методы, эмулирующие работу метода обратного распространения ошибки за счёт особого подбора параметров STDP[1646], [1647], [1648]. Эти методы для ряда задач демонстрируют эффективность, сходную с эффективностью метода обратного распространения ошибки[1649], [1650], [1651]. 5.3.7 Нейроморфные системы типа I. Перспективы Одним из потенциальных преимуществ импульсных нейронных сетей является, по всей видимости, то, что они меньше подвержены так называемой проблеме катастрофической интерференции [catastrophic interference] или, как её ещё называют, проблеме катастрофического забывания [catastrophic forgetting][1652].
Суть этой проблемы заключается в том, что искусственные нейронные сети, обученные на некоторой выборке, имеют склонность внезапно полностью забывать ранее изученную информацию при попытке «доучить» их на другой выборке, содержащей прецеденты, с точки зрения человека отличающиеся от прецедентов в оригинальной выборке. Это существенно осложняет перенос обучения (знаний) [transfer learning] между близкими областями и создаёт проблемы при дообучении моделей [fine-tuning, дословно — тонкая настройка]. Впрочем, для классических нейронных сетей в настоящее время существует несколько рабочих рецептов, предназначенных для борьбы с этой проблемой (мы поговорим о них позже). вернуться Levy W. B., Steward O. (1983). Temporal contiguity requirements for long-term associative potentiation/depression in the hippocampus / Neuroscience, Vol. 8, Iss. 4, April 1983, pp. 799—808 // https://doi.org/10.1016/0306-4522(83)90011-8 вернуться Artola A., Brocher S., Singer W. (1990). Different voltage-dependent thresholds for inducing long-term depression and long-term potentiation in slices of rat visual cortex / Nature, Vol. 347, pp. 69—72 // https://doi.org/10.1038/347069a0 вернуться Markram H., Gerstner W., Sjöström P. J. (2011). A history of spike-timing-dependent plasticity / Frontiers in synaptic neuroscience, 3, 4 // https://doi.org/10.3389/fnsyn.2011.00004 вернуться Debanne D., Gahwiler B. H., Thompson S. M. (1994). Asynchronous pre- and postsynaptic activity induces associative long-term depression in area CA1 of the rat hippocampus in vitro / Proceedings of the National Academy of Sciences of the United States of America, Vol. 91 (3), pp. 1148—1152 // https://doi.org/10.1073/pnas.91.3.1148 вернуться Malinow R. (1991). Transmission between pairs of hippocampal slice neurons: quantal levels, oscillations, and LTP / Science, Vol. 252, Iss. 5006, pp. 722—724 // https://doi.org/10.1126/science.1850871 вернуться Verstraelen P., Van Dyck M., Verschuuren M., Kashikar N. D., Nuydens R., Timmermans J.-P., De Vos W. H. (2018). Image-Based Profiling of Synaptic Connectivity in Primary Neuronal Cell Culture / Frontiers in Neuroscience, 26 June 2018 // https://doi.org/10.3389/fnins.2018.00389 вернуться Danielson E., Lee S. H. (2014). SynPAnal: Software for Rapid Quantification of the Density and Intensity of Protein Puncta from Fluorescence Microscopy Images of Neurons / PLoS One, Vol. 9 (12), e115298 // https://doi.org/10.1371/journal.pone.0115298 вернуться Kashiwagi Y., Higashi T., Obashi K., Sato Y., Komiyama N. H., Grant S. G. N., Okabe S. (2019). Computational geometry analysis of dendritic spines by structured illumination microscopy / Nature Communications, Vol. 10, Article number: 1285 // https://doi.org/10.1038/s41467-019-09337-0 вернуться Markram H., Sakmann B. (1995). Action potentials propogating back into dendrites triggers changes in efficacy of single-axon synapses between layer V pyramidal cells / Society for Neuroscience abstracts, Vol. 21. вернуться Markram H., Gerstner W., Sjöström P. J. (2011). A history of spike-timing-dependent plasticity / Frontiers in synaptic neuroscience, 3, 4 // https://doi.org/10.3389/fnsyn.2011.00004 вернуться Song S., Miller K. D., Abbott L. F. (2000). Competitive Hebbian learning through spike-timing-dependent synaptic plasticity / Nature Neuroscience. Vol. 3, pp. 919—926 // https://doi.org/10.1038/78829 вернуться Markram H., Gerstner W., Sjöström P. J. (2011). A history of spike-timing-dependent plasticity / Frontiers in synaptic neuroscience, 3, 4 // https://doi.org/10.3389/fnsyn.2011.00004 вернуться Izhikevich E. M. (2007). Solving the distal reward problem through linkage of STDP and dopamine signaling / Cerebral Cortex, Vol. 17, pp. 2443—2452 // https://doi.org/10.1093/cercor/bhl152 вернуться Frémaux N., Gerstner W. (2016). Neuromodulated spike-timing-dependent plasticity, and theory of three-factor learning rules / Frontiers in Neural Circuits, Vol. 9 // https://doi.org/10.3389/fncir.2015.00085 вернуться Tavanaei A., Maida A. (2019). BP-STDP: Approximating backpropagation using spike timing dependent plasticity / Neurocomputing, Vol. 330, pp. 39—47 // https://doi.org/10.1016/j.neucom.2018.11.014 вернуться Bengio Y., Mesnard T., Fischer A., Zhang S., Wu Y. (2017). STDP-compatible approximation of backpropagation in an energy-based model / Neural computation, Vol. 29, Iss. 3, pp. 555—577 // https://doi.org/10.1162/NECO_a_00934 вернуться Millidge B., Tschantz A., Buckley C. L. (2020). Predictive coding approximates backprop along arbitrary computation graphs // https://arxiv.org/abs/2006.04182 вернуться Mozafari M., Ganjtabesh M., Nowzari-Dalini A., Thorpe S. J., Masquelier T. (2019). Bio-Inspired Digit Recognition UsingSpike-Timing-Dependent Plasticity (STDP) and Reward-Modulated STDP in Deep Convolutional Networks / Pattern Recognition, Vol. 94, pp. 87—95 // https://doi.org/10.1016/j.patcog.2019.05.015 вернуться Lee C., Panda P., Srinivasan G., Roy K. (2018). Training Deep Spiking Convolutional Neural Networks With STDP-Based Unsupervised Pre-training Followed by Supervised Fine-Tuning / Frontiers in Neuroscience, Vol. 12, 2018 // https://doi.org/10.3389/fnins.2018.00435 вернуться Mozafari M., Kheradpisheh S. R., Masquelier T., Nowzari-Dalini A., Ganjtabesh M. (2018). First-Spike-Based Visual Categorization Using Reward-Modulated STDP / IEEE Transactions on Neural Networks and Learning Systems, Vol. 29, No. 12, pp. 6178—6190 // https://doi.org/10.1109/TNNLS.2018.2826721 вернуться Vaila R., Chiasson J., Saxena V. (2019). Deep Convolutional Spiking Neural Networks for Image Classification // https://arxiv.org/abs/1903.12272 |