Важность проблемы топологии нейронных сетей осознавалась как Розенблаттом, который посвятил этой теме немалую долю текста «Принципов нейродинамики», так и Минским в его анализе способности нейронных сетей вычислять предикаты чётности и связности. Каким бы мощным инструментом ни был метод обратного распространения ошибки, задача обучения больших полносвязных сетей довольно быстро становится неподъёмной по мере увеличения количества их нейронов. Да и естественные нейронные сети в своей топологии явно далеки от полносвязных: каждый нейрон человеческого мозга связан лишь с несколькими тысячами из почти сотни миллиардов других нейронов. В своих поисках эффективных архитектур искусственных сетей Розенблатт, вполне ожидаемо, обратился к данным нейрофизиологии.
В «Принципах нейродинамики» содержатся ссылки на наиболее передовые для начала 1960-х исследования в области нейрофизиологии. Здесь мы находим отсылки к работам Роджера Сперри; Летвина, Мак-Каллока, Питтса и Матураны (та самая работа про глаз и мозг лягушки); Стюарта Сазерленда (исследование зрения осьминогов) и замечательным результатам, полученным Торстеном Визелем и Дэвидом Хьюбелом в процессе изучения зрительной коры кошек[1449]. Примечательно, что Сперри, Визель и Хьюбел в 1981 г. разделят Нобелевскую премию в области физиологии и медицины.
Существует мнение, что Розенблатт вплотную приблизился к идее создания свёрточных нейронных сетей[1450]. Как показывают его статьи[1451], [1452], он пытался построить нейронную сеть, которая бы моделировала работу зрительного аппарата живого существа. Эксперименты Розенблатта с элементарными перцептронами явственно демонстрировали, что полносвязная или связанная случайным образом сеть обладает низкой эффективностью, то есть неспособна учиться достаточно быстро. Розенблатт искал механизмы, позволяющие сократить число параметров модели, а также объяснить существование механизма долгосрочной памяти. Эти теоретические размышления, а также результаты опытов по переносу памяти у крыс убедили его в том, что в мозге существует механизм, позволяющий передавать информацию о величинах синаптических весов из одной области в другую. Розенблатт предположил, что это может происходить химическим путём, но в итоге наука нашла другие объяснения (хотя и отдалённо похожие на идеи Розенблатта). Этому как раз способствовали результаты, полученные Хьюбелом и Визелем.
5.2.4.1 Роль котиков в развитии нейронных сетей
Хьюбел и Визель занимались изучением первичной зрительной коры (V1), также называемой «полем Бродмана 17» или же «стриарной корой». Последнее название происходит от латинского слова stria, то есть «полоса», и отражает тот факт, что здесь отчётливо видна полоска Дженнари (наружная полоска Байярже), образованная конечными отделами покрытых миелиновой оболочкой аксонов, отходящих от нейронов латерального коленчатого тела и заканчивающихся в IV слое серого вещества. Выучив все эти термины, вы сможете без особого труда подавлять в споре недостаточно уверенных в себе оппонентов.
Именно в зрительную зону V1 попадают нервные импульсы нейронов зрительного нерва, генерируемые сетчаткой. Все остальные чувствительные к зрительным сигналам участки мозга связаны с глазами строго через зону V1. У млекопитающих зона V1 занимает задний полюс затылочной доли каждого полушария. Это наиболее простая и по происхождению более «древняя» из зон коры головного мозга, связанных со зрением.
Но перед рассмотрением эксперимента Хьюбела и Визеля надо обсудить важные особенности строения коры головного мозга.
Кора головного мозга человека представляет собой плотно уложенный складками слой нервной ткани толщиной около двух — четырёх миллиметров[1453]. Если развернуть этот слой, то площадь коры одного полушария составит порядка 1000–1400 квадратных сантиметров[1454]. Под микроскопом в ткани коры можно разглядеть множество нейронов — примерно 100 000 на квадратный миллиметр.
Клеточные тела образуют несколько слоёв, в каждом из которых плотность их размещения практически постоянна, но более плотные слои перемежаются менее плотными. Ещё в конце XIX в. учёным удалось установить, что кора подразделяется на области, выполняющие различные функции (локализация функций). Обширным источником знаний о функциональных зонах коры стали клинические данные. Врачи неоднократно отмечали, что черепно-мозговые травмы, в зависимости от их локализации, могут вызвать слепоту, паралич, онемение или потерю речи, при этом слепота может быть как полной, так и ограниченной некоторой частью зрительного поля, а онемение может затрагивать как всю конечность, так и только несколько пальцев. Изучение взаимосвязей между местом повреждения коры и участком проявления на теле позволило со временем составить общую карту коры, выделив тем самым зрительную, слуховую, соматосенсорную (отвечающую за ощущения тела), речевую и моторную кору.
Тщательный анализ коры с помощью микроскопа и улучшенных способов окраски показал, что, несмотря на относительную однородность, в коре существуют структурные различия, хорошо коррелирующие с картой локализации функций. Дополнительные подтверждения этого были получены в ходе изучения электрической активности мозга, возникающей при стимуляции организма при помощи прикосновений, звуков или световых раздражителей. Сходным образом, наблюдая за движениями тела в ответ на электрическую стимуляцию коры, удалось также составить и карту двигательных зон. Это систематическое картирование коры вскоре привело к фундаментальному выводу: в большинстве случаев сенсорные и моторные зоны мозга представляли собой «проекции» соответствующих областей тела. Стало ясно, что тело сопоставлено соматосенсорной и моторной областям коры, при этом полю зрения соответствует первичная зрительная кора площадью около 15 квадратных сантиметров. В первичной зрительной коре карта достаточно проста, поскольку не содержит НИ ЕДИНОГО РАЗРЫВА, за исключением примечательного разделения поля зрения посередине, причём левая его половина проецируется на кору правого полушария головного мозга, а правая — на кору левого полушария.
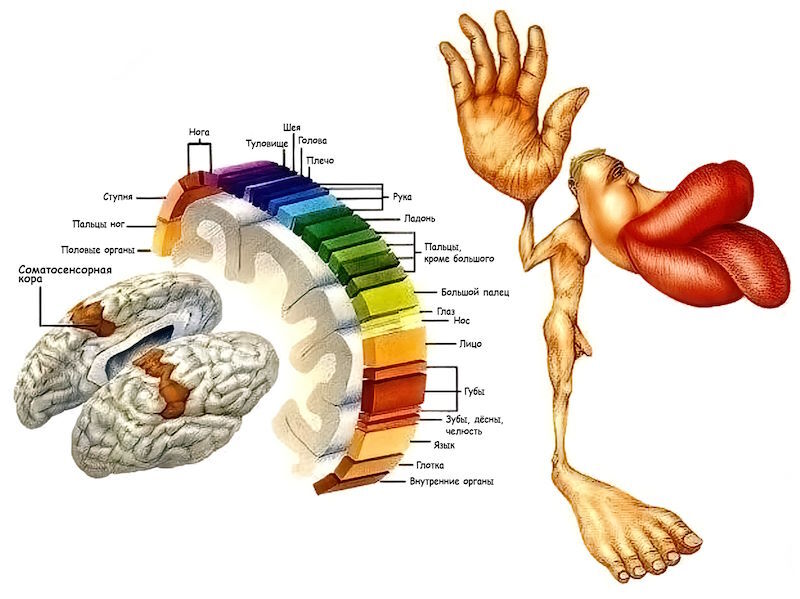
Важной особенностью корковых (также говорят «кортикальных», от лат. corticalis — кора) карт является их переменный масштаб: одна и та же площадь поверхности тела, в зависимости от конкретного участка, может отображаться в различные по площади участки коры. Например, квадратному миллиметру поверхности пальцев, губ или языка соответствует бóльшая площадь коры, чем квадратному миллиметру поверхности ягодиц или спины, а квадратный миллиметр центральной части сетчатки глаза представлен в коре мозга примерно в 35 раз детальнее, чем квадратный миллиметр её периферической части.
Во многих учебниках по нейрофизиологии можно обнаружить изображение удивительного существа — соматосенсорного гомункулуса. В основу гомункулуса положена фигурка человека, но размеры разных анатомических областей его тела выполнены в масштабе, пропорциональном числу находящихся в них рецепторов и, соответственно, площади, приходящейся на эти анатомические области зоны соматосенсорной коры.
Рис. 102. Соматосенсорный гомункулус
Эта особенность кортикальных карт была задокументирована в начале 1940 гг. — в пионерских исследованиях Клинтона Вулси[1455] и Эдгара Эдриана[1456], в которых было отмечено, что частям тела, обладающим наибольшей чувствительностью, соответствуют наибольшие по площади зоны кортикальных проекций[1457].