Весьма похожие между собой строением мозга, но столь разные внешне, шу-ю и ромундина (с зубастыми челюстями и парными грудными плавниками) вместе с другими панцирными бесчелюстными позвоночными и пластинокожими рыбами образуют почти непрерывный ряд переходных форм. Что-то менялось медленно, например мозг, что-то быстро – так, видимо, возникли челюсти и парные плавники. Сравнивая всех этих животных между собой, можно убедиться, что многие представления о ранней эволюции челюстноротых, сложившиеся в XIX в. и укоренившиеся на страницах учебников и зоологических сводок, – казалось бы, весьма архаичные и даже забавные – все равно в чем-то верны. Да, челюсти – это не сложившаяся пополам древняя жаберная дуга, а совершенно новый орган, как уже сказано в предыдущей главе. И парные многолучевые плавники не ведут свое начало от разросшихся шипастых жаберных перегородок, боковых складок, с двух сторон охватывавших туловище, или парных шипов.
Наоборот, плечевой пояс, вероятно, вел свое начало от шестой жаберной дуги и первоначально служил для прикрепления мускулов, открывавших пасть. Лишь позднее этот блок стал использоваться для управления грудными плавниками. Этот переход наблюдался у колымасписа (Kolymaspis) – близкого родственника ромундины, т. е. акантоторакса со звездчатым черепом, плававшего в раннедевонскую эпоху на востоке Сибири. Верхняя часть дуги у него играла роль черепно-плечевого сустава, а жаберная щель отделяла плечевой пояс от головы.
У многих бесчелюстных парные плавники уже появились, причем прежде боковых шипов или жаберных перегородок с лучами, а складки, которые могли бы распасться на отдельные плавники, окаймляли туловище у единичных видов. Зато на примере ископаемых мы видим, что парные конечности могли возникнуть почти в любом месте – в грудном отделе (остеостраки, телодонты и рыбы), в брюшном (челюстноротые), и даже анальные плавники могли удваиваться (некоторые анаспиды и рыбы). И скорее такие парные плавники срастались в протяженные продольные складки, а не наоборот.
Все зависит от работы определенных генов и целых регуляторных участков генома, которые способны переключаться на другие части тела или удваиваться, вызывая развитие парных конечностей там, где их раньше не было. В наиболее далеко зашедших случаях получаются плавниковые складки или, скажем, раздвоенный хвостовой плавник, причем не с верхней и нижней лопастями, а с двумя отдельными двулопастными «хвостами». Для дикого животного это верная смерть: попробуй плавать, когда правая часть реально не знает, что делает левая. А вот в домашних условиях золотых рыбок с ярко выраженным раздвоением личности в области хвоста японцы разводят уже более четырех столетий. Конечно, столь экзотичные даже среди обитателей аквариумов существа не могли не заинтересовать японских ученых. Анатомия этих удивительных карасей была скрупулезно описана еще в конце XIX в. Исследования показали, что у них произошло удвоение мускульных и скелетных элементов хвоста, включая плавниковые лучи. Недавно генетики Страны восходящего солнца выяснили, что одна-единственная мутация в одном из двух генов chordin приводит к новому типу заложения конечностей в период эмбрионального развития и вместо непарного плавника вырастает раздвоенный.
Тот же генный комплекс в какой-то степени влияет на формирование жаберных дуг, но не челюстей и не гиоидной дуги, тесно связанных с нервным гребнем. У эволюции очень бережное отношение ко всем прежним наработкам. Все можно пустить в дело, и еще не раз, на совершенно ином уровне и в новых группах организмов, создавая то парные плавники с лучами, то лапы с пальцами. Это отнюдь не означает, что при переходе от бесчелюстных к челюстноротым сработал именно этот генный механизм, и всего единожды: просто фундаментальные, как нам кажется, вопросы в эволюции могут решаться довольно просто…
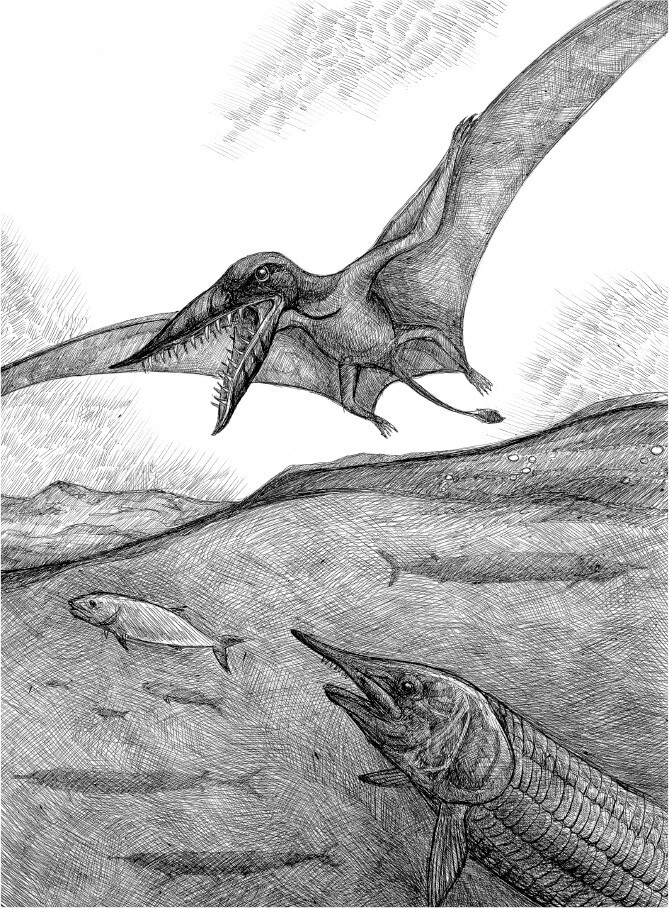
В спокойных околорифовых водах над белыми известковыми илами, населенными одиночными роговидными кораллами, двустворчатыми брахиоподами и морскими лилиями, прицепившимися к раковинам длинными членистыми стебельками, медленно маневрировала стайка рыбообразных существ. Даже удивительно, как эти животные, облаченные в конический панцирь, заканчивающийся огромным спинным шипом, передвигались, поводя лишь хвостовым плавником. Их глаза в поисках свободных от раковин проплешин, где могли затаиться беззащитные черви, смотрели вниз и не заметили, как из рифовой ниши выскочила рыбка, раза в два уступавшая им размером. Рыбка, тоже в панцире, но облегченной конструкции, проворно помогая себе грудными плавничками, вдруг оказалась под брюхом у одного из, казалось бы, лишенных слабых мест панцирников. Она решительно вцепилась в него мелкими, но острыми зубными пластинками, высмотрев едва заметную брешь между щитками. Зеленовато-голубая вода побурела, и вдруг на запах крови из фиолетовой бездны, куда круто обрывалась рифовая отмель, всплыла колючая рыбина с пастью, полной пилообразных зубов. В этой пасти мгновенно оказались и маленький хищник, и его жертва, усеяв дно кусками панциря.
Так могло случиться примерно 415 млн лет назад. Пластинокожие рыбы, даже такие небольшие, как ромундина, действительно охотились на бесчелюстных гетеростраков, подобных описанному здесь митраспису (Mitraspis), около 30 см длиной. А вершину пищевой пирамиды в раннедевонских морях, в том числе и Лавруссии, где находился ныне заполярный остров Принца Уэльского, занимали акантоды (Acanthodii, от греч. ακανθα – «колючка»), родственные акулам и похожие на них мощными плавниками и зубными рядами, выгнутыми как подкова. На обломках панцирей бесчелюстных следы укусов не редкость. Их число резко возросло в девонском периоде с появлением рыб, оснащенных челюстями. Получается, что челюстноротые если и не съели всех панцирных бесчелюстных, то очень многих понадкусывали.
Глава 6
Пронзающий птеродактилей. Аспидоринх
После всевозможных палеозойских панцирников лучеперые костистые рыбы могут показаться кому-то заурядными. Их мы видим везде: в морях, реках, небольших прудиках и ручьях, фонтанах и аквариумах, не говоря уж о прилавках рынков, магазинов и меню ресторанов, даже если это не «фиш-энд-чипс». Но и эти рыбы пережили несколько важных этапов эволюции: палеозойские были мало похожи на мезозойских, а окружающие нас кайнозойские и вовсе в большинстве другие. Расцвет последних совпал с эоценовой – миоценовой эпохами и бурным развитием коралловых рифов, приютивших почти половину их разнообразия и многоцветия: от рыб-клоунов, ласточек, хирургов и попугаев до скорпен, мурен и груперов.
От былого палеозойско-мезозойского великолепия почти ничего не осталось: хрящевых и костных ганоидов, вместе взятых, и полусотня видов с трудом наберется. Почти все они населяют пресные водоемы Северного полушария или нерестятся там. Многоперы уцелели на севере Африки, ильная и каймановые рыбы – только в Северной Америке, и лишь осетровые обитают по всей Северной Америке и Евразии. И те на глазах исчезают, как недавно признанный окончательно вымершим псефур из реки Янцзы. А ведь рыба была немаленькая – до 7 м длиной, с учетом огромного мечевидного рыла. Осетровым вообще не повезло: единственными достоверными жертвами чиксулубского метеоритного взрыва, завершившего мезозойскую эру, оказались именно они, попав под сокрушительный удар сейши (мощной стоячей волны). А дождь из капель каменного расплава обернулся в воде твердыми сферулами – тектитами, и они забили рыбам жабры. Не спасло даже то, что стая обитала в устье реки Танис, впадавшей в Западное внутреннее море на территории современного штата Северная Дакота, – очень далеко от места «жесткой посадки» небесного тела (полуостров Юкатан). Случилось это на исходе северной весны. Что вовсе не красивая фраза, а факт, установленный по сезонным изменениям в соотношении стабильных изотопов кислорода и углерода, плотности остеоцитов и ширины линий нарастания в зубной кости и грудном плавнике погибших осетров и веслоносов.