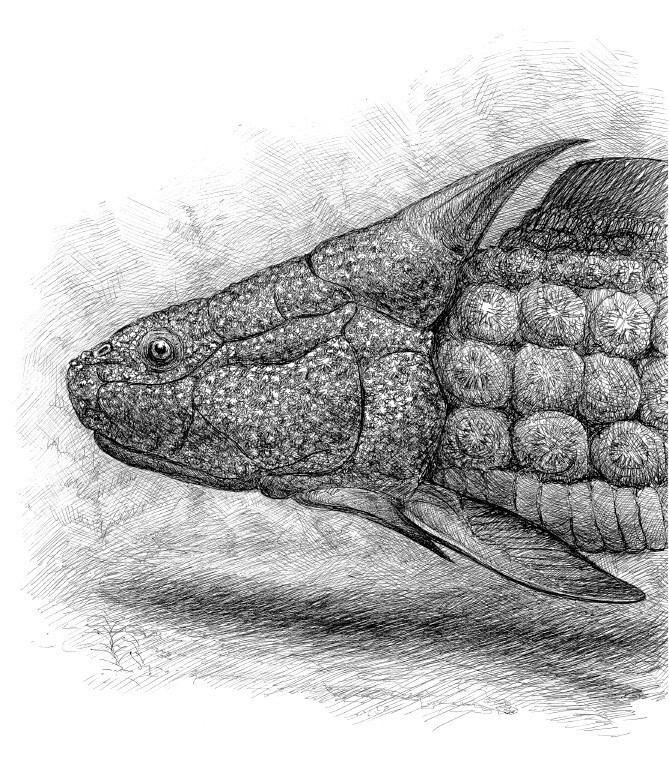
Мы никогда не увидим, из каких именно клеток состояли важные черепно-лицевые нервы шу-ю и его современников. Однако раздвоенный носовой мешок указывает на появление двух дополнительных боковых плакод. И, значит, клетки нервного гребня у этого бесчелюстного уже устремились вперед, образуя нервные узлы нового типа, а попутно и ткани, где закладывались челюсти. А в том, что челюсти у рыб в какой-то степени уподобились жаберным дугам, нет ничего удивительного: новый орган развивался в прямой зависимости от уже имевшегося генного комплекса.
На некоторых образцах шу-ю можно увидеть детали кровеносной системы. В них он тоже ближе к челюстноротым, чем к своим названым родственникам – вымершим остеостракам и еще живым миногам. Скажем, спинная аорта, когда-то несшая насыщенную кислородом кровь от жаберных лепестков ко всем органам тела, у галеаспид была парной и проходила через хрящевое основание черепа. И укрупненные боковые вены головы, направлявшие отработанную кровь к предсердию, занимали то же место, где у челюстноротых расположились яремные вены. Появились глазнично-носовые артерии и вены, что опять же связано с раздвоением носового мешка. А вот разветвленная и расходившаяся к краям головотуловищного щита подкожная сеть тончайших капилляров, питаемых поверхностной артерией, в мире позвоночных, пожалуй, аналогов не имеет. Не исключено, что эта сеть обслуживала канальцы, подобные электрочувствительным органам остеостраков.
Шу-ю помог раскрыть и тайну происхождения еще одного важного органа позвоночных – барабанной полости среднего уха и связанной с ней евстахиевой трубы. Предполагалось, что это внутричерепное пространство образуется на основе эмбрионального подъязычно-нижнечелюстного (жаберного) кармана. А карман является рудиментом брызгальца, которое когда-то было полноценной жаберной щелью, расположенной между самостоятельными челюстной и гиоидной дугами. (Брызгальце как функциональная полость сохранилось у многих хрящевых и некоторых «древних» костных рыб, таких как многоперы и осетровые, а также у форм, переходных между рыбами и наземными четвероногими, и даже у первых рептилий.) Вот только найти эту гипотетическую часть жаберного скелета ни у кого из давно вымерших рыб не удавалось. Вероятно, потому, что такая дуга перестала выполнять свои изначальные функции уже ко времени появления первых челюстноротых. Ее верхний элемент (гиомандибула) превратился в подвеску для нижней челюсти, а затем (у наземных четвероногих) крайний «кусочек» этого элемента стал одной из слуховых косточек (стременем); нижняя часть стала подъязычной костью. А вот у бесчелюстного шу-ю полноценная гиоидная жаберная дуга и соответствующая щель еще были!
Прочие части тела у шу-ю не сохранились, но как они выглядели, подсказывают остатки других галеаспидов. Например, у туцзяасписа (Tujiaaspis, от «туцзя» – имени народа, населяющего центральные провинции Китая, в том числе Хунань, откуда описано это ископаемое) все туловище, которое было значительно у́же головного щита и лишь ненамного длиннее, покрывали плотные ряды ромбовидных чешуй (16 штук на каждый квадратный миллиметр). Придатки его тела выглядели несколько необычно: три почти треугольных спинных плавника с отогнутыми назад вершинами, серповидная хвостовая лопасть и две длинные боковые складки, тянувшиеся от жаберных отверстий до самого хвоста и состоявшие из скобообразных скелетных элементов. Эти жесткие скобы успешно рассекали водную толщу, а вихревые течения, начинавшиеся у задних выемок головного щита, направляли поток под брюхо и тем самым увеличивали подъемную силу. Как и все бесчелюстные панцирники, в преодолении лобового сопротивления туцзяаспис больше полагался на аэродинамическую форму тела, чем на мускульные усилия плавников.
Внешне похожие на прочих панцирных бесчелюстных, галеаспиды, особенно шу-ю, воспринимали мир как челюстноротые. Шу-ю не успела стать рыбой, но вполне оправдывала свое китайское имя «ранняя рыба».
Глава 5
Многозвездный броненосец. Ромундина
Итак, пути эволюции круглоротых и прочих бесчелюстных разошлись очень рано – даже раньше, чем от «прочих» отделились зубастые конодонтоносцы. Некоторые специалисты, правда, не исключают, что предшественниками миног и миксин являлись анаспиды – самые беспанцирные из панцирных бесчелюстных, причем с веретеновидным (рыбьим) телом. Но и те имели покров из плотно уложенных продолговатых трехслойных чешуй (энамелоид – дентин – бесклеточная кость), парные плавники и сложный кишечник. Потому другие ученые сближают их с челюстноротыми. В этом случае анаспидам понадобилось бы вновь собрать чешуи в панцирь, поскольку наиболее примитивными настоящими рыбами стали пластинокожие (Placodermi), господствовавшие в морях всего света с раннесилурийской по позднедевонскую эпоху (435–360 млн лет назад).
Самые ранние из них очень напоминали бесчелюстных панцирников: такой же уплощенный головной щит с отходившими назад шипами и небольшое чешуйчатое тельце, заканчивавшееся эпицеркальным хвостом. Все вместе – 3 см длиной. Но они обзавелись минерализованными позвонками, составным жаберным скелетом, плавниковыми лучами и, что самое главное, челюстями и связанной с ними мощной мускулатурой. Челюсти позволили им превратиться в девонском периоде в первых позвоночных гигантов – дунклеостеев (Dunkleosteus), до 4 м длиной, превзойдя крупнейших активных беспозвоночных хищников – ракоскорпионов (до 2,5 м). Одной из веских причин резкого роста рыбьих размеров – в 10 раз по сравнению с силурийскими предшественниками – стало долгожданное повышение уровня кислорода до современного. Без достаточного количества этого основного и дармового энергетика не просто вволю не надышишься, но, главное, не вымахаешь до крупных размеров, будучи активным хищником с массивным скелетом.
Почти до конца девонского периода бесчелюстные панцирники не сходили с эволюционной сцены, а когда ушли – мир стал принадлежать челюстноротым (см. рис. 4.2). Ныне из примерно 58 000 видов позвоночных более 99 % составляют представители этой группы.
Это неудивительно, особенно для обитателей водного мира. Конструкция челюстного аппарата у большинства рыб (а также морских черепах, тюленей, китов, фламинго и уток) такова, что достаточно широко раскрыть рот, и пища влетит туда сама, увлекаемая потоком воды из-за перепада давления во внешней среде и ротоглоточной полости. На эти физические законы уповают многие засадные хищники – от позднепалеозойских акул и «лягушкоящеров» до современных щук, сомов и гигантских саламандр. Позвоночное без челюстей обречено всю жизнь плавать в поисках пищи или стать почти сидячим мелким фильтратором, как пескоройки. Но у древнейших (силурийских и девонских) рыб, судя по челюстям, этот механизм захвата пищи срабатывал резко и, главное, «аккуратно, но сильно». Для этого лучше всего подходили спрямленные челюсти с центром масс, смещенным ближе к месту сочленения. Челюстноротые изначально были нацелены на фастфуд в прямом смысле этого слова. Они «позиционировали» себя как сноровистые хищники. (Другие, специализированные, варианты появились позднее.)
Ротовой аппарат пластинокожих рыб еще не приобрел всех деталей настоящей помпы, что не помешало им стать в девонском периоде самой многочисленной и достаточно разнообразной группой. Особенно выделялись огромные хищные артродиры (Arthrodira, от греч. αρθροω – «сочленять» и δερη – «шея»), к которым принадлежали и дунклеостей, и другие гиганты. На современных – обычно веретеновидных и сжатых с боков – рыб они ничуть не походили. За огромной головой, ширина и высота которой были примерно равны, следовало тело, длина которого лишь немногим превышала таковую головной части.
Ворочать тяжелой головой и отдельно плечевым поясом с плавниками им помогала целая система брюшных мышечных блоков. Она состояла из пучков упругих и прочных поперечнополосатых мышечных волокон и была сравнима с мускулатурой наземных позвоночных, но не рыб. Челюстной аппарат дунклеостея работал как четырехзвенный шарнирный механизм, где роль шарниров выполняли не только верхняя и нижняя челюсти, но и мозговая коробка черепа, и даже плечевой пояс (рис. 5.1). Двигали шарнирами мощные мышцы. Пасть открывалась не более чем за 130 микросекунд: не так быстро, чтобы создать водоворот, мгновенно увлекающий добычу, но достаточно для того, чтобы вызвать всасывающий эффект, подобно разинутому рту акулы-няньки. То, что не влетало сразу из-за неподходящего размера, кромсалось на части. Расчеты силы смыкания похожих на две острозаточенные секиры челюстных пластин у дунклеостея показывают, что эти лезвия срабатывали быстрее гильотины, круша добычу с силой от 4400 до 5300 Н. (Пятнистая гиена для разгрызания костей прилагает меньше усилий.) Когда с возрастом зубные выросты стачивались, гиганты еще какое-то время продолжали жить, давя пищу уцелевшими пластинами.