Таким образом, из 13 известных для современного Алтая видов Chiroptera, на исследованной территории к настоящему моменту установлено 7 видов. Большинство перечисленных летучих мышей в своем распространении связаны с широколиственными и/или хвойно-широколиственными лесами. Обитателем речных пойм лесной и лесостепной зон Европы, Южной Сибири, Китая и Японии является водяная ночница.
Суммируя приведенные наблюдения, можно констатировать, что в долине Ануя в настоящее время преобладают млекопитающие таежного комплекса. В окрестностях Денисовой пещеры он имеет господствующее положение как по занимаемой площади, так и по численности слагающих его видов. Меньшее значение имеют луговые сообщества. Степные ассоциации в настоящее время отсутствуют в этом районе. Однако их отдельные представители вкраплены в сообщества луговых и агрокультурных ценозов.
Такие достаточно подробные данные по видовому составу современной фауны млекопитающих долины Ануя, относительной численности отдельных видов и их приуроченности к определенным биоценозам стали сравнительной основой для изучения тафономических процессов в современных условия Северо-Западного Алтая. На протяжении последних 10 лет на стационаре «Денисова пещера» проводится сбор погадок хищных птиц экскрементов хищных млекопитающих, анализируются материалы Современных осадков многочисленных ниш, гротов, скальных щелей. Сравнение полученных результатов с данными учетов ловушками позволяет понять как изменяется видовой состав и численное соотношение фаунистического комплекса на пути от биоценоза к танатоценозу. В свою очередь это станет ключом к расшифровке информации по мелким млекопитающим из отложений палеолитических памятников.
Анализ 140 погадок коршуна показал, что первое место в его добыче занимает обыкновенная полевка — 35,5 %. Численность остальных видов значительно меньше. Длиннохвостый суслик и крот составляют около 4 % каждый. Землеройки, преимущественно Sorex araneusL., - 7 %. На долю рыжей полевки Cleîhrionomysприходится лишь 3,5 % от общего количества остатков. Около 5 % составляет цокор. Мыши и хомячок не превышают каждый 0,7 %. В погадках обнаружены остатки серой крысы Rattus norvegicusBerkenhout — 2,1 %, водяной полевки Arvicola terrestrisL. - 0,7 % и обыкновенного хомяка Cricetus cricetusL. - 0,7 %. Три последних вида дополняют список современной фауны долины Ануя, установленный по учетам ловушками. Серая крыса связана с постройками человека. Водяная полевка, судя по кормовым столикам, обитает в пойме Ануя и Каракола повсеместно. Однако она везде малочисленна, что подтверждают и материалы погадок. Обыкновенный хомяк, по аналогии с другими районами Алтая, вероятно, приурочен к сельскохозяйственным угодьям.
Приведенные материалы позволяют сделать два существенных вывода. Во-первых, список современной фауны долины Ануя должен быть дополнен водяной полевкой, обыкновенным хомяком и серой крысой. Во-вторых, в погадках хищников количественное соотношение видов мелких млекопитающих не вполне соответствует их количественному соотношению в природе, что было показано ранее [ Громов1957; Kowalski1990]. Хищные птицы явно охотятся избирательно, предпочитая формы открытых биотопов. В их погадках концентрируются виды, обитающие на лугах, опушках, полянах. Доля видов зверьков, живущих под пологом леса и в зарослях кустарников, занижена по сравнению с их численностью в природе. Это необходимо принимать во внимание при реконструкции ландшафтов по палеофаунистическим данным.
3.2. Ископаемая фауна млекопитающих бассейна Ануя по материалам палеолитических памятников
3.2.1. Палеолитический памятник Денисова пещера
Денисова пещеры расположена в Солонешенском районе Алтайского края в 6 км северо-западнее с. Черный Ануй ив 1,8 км ниже устья р. Каракол по правому борту каньона р. Ануй. Вход в пещеру имеет юго-западную экспозицию и расположен в 28 м над урезом р. Ануй. Денисова пещера представляет собой карстовую полость в силурийских известняках. Высота центральной камеры от поверхности голоценовых отложений около 10 м. Глубина скального основания в центральной части около 6 м. Детально профиль пещеры и прилегающих галерей описан ранее [ Деревянко, Молодин1994]. Мощность голоценовых отложений незначительно варьирует по простиранию и в среднем равна 1,5 м. Мощность вскрытых плейстоценовых отложений составляет около 4,5 м. Получены датировки радиоуглеродным (С 14) и радио-термолюминисцентным (РТЛ) методами плейстоценовой части разреза Денисовой пещеры, которые приведены в таблице 1.
Таблица 1. Датировки плейстоценовых отложений Денисовой пещеры [ Деревянко и др.1998].
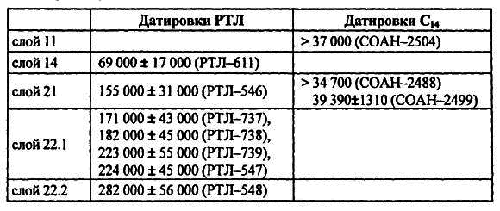
Приведенные датировки сами по себе имеют значительный разброс. Более того, они не соответствуют результатам полученным на основании изучения состава фауны мелких млекопитающих и эволюционного уровня отдельных ее компонентов. Судя по этим результатам, вся плейстоценовая толща Денисовой пещеры укладывается в интервал позднего плейстоцена. При этом, нижняя часть осадков (слои 22.1; 22.2; 22.3) соответствует первой половине позднего плейстоцена, т. е. казанцевскому межледниковью. Слои 20-9 отвечают второй половине позднего плейстоцена, т. е. ермаковскому, каргинскому и сартанскому времени. Если принять такое датирование отложений, то каждый горизонт 22 слоя и каждое литостратиграфическое подразделение в интервале 21-9 слой соответствует временному интервалу 5–6 тыс. лет. Если принять РТЛ датировки, то возрастной интервал каждого слоя плейстоценовых отложений Денисовой пещеры приблизительно 10–15 тыс. лет. Возрастной интервал для литостратиграфических подразделений голоценовой толщи около 1 тыс. лет.
Ниже приведены результаты определения сборов 1994 и 1995 годов. Сборы 1994 г.: квадраты Е-6, Е-7, Б-8 и Б-9 центрального зала (сектор 4) Денисовой пещеры по линии субквадратов «а» и «в». Сборы 1995 г.: квадраты Е-5 (субквадраты а, б, в, г), Е-6, Е-7, Е-8 (субквадраты б, г), Е-9 (субквадраты б, г) центрального зала (сектор 4) Денисовой пещеры. Заключения по этим двум параллельно расположенным профилям сделаны отдельно для каждой из коллекций Это обусловлено рядом причин. Дело в том, что отложения Денисовой пещеры имеют очень сложное пространственное залегание. Ряд слоев имеет линзовидный характер, а некоторые — сильно перемяты и смещены за счет конвективной неустойчивости грунтов. От квадрата к квадрату меняется мощность слоев, нарушается их последовательность за счет выклинивания некоторых из них. Неравномерна концентрация костного материала по площади. Все это придает высокую тафономическую индивидуальность каждому квадрату раскопа. Фактически каждый из них неповторим по условиям осадконакопления и набору остатков млекопитающих. При столь сложной тафономической ситуации необходима дифференцированная оценка получаемых результатов. Только так можно отделить случайные флуктуации таксономического разнообразия от изменений, отражающих истинную динамику состава фауны.
3.2.1.1. Результаты изучения сборов 1994 г. сектора 4 Денисовой пещеры
Общее количество определенных костных остатков в коллекции 1994 г. составляет 6353 экземпляра. В среднем каждый слой охарактеризован приблизительно 200 определенными остатками. В целом верхние слои охарактеризованы менее надежно, чем нижние. Общий список таксонов, установленных для плейстоценовых слоев Денисовой пещеры, включает более 40 названий. Их количественное соотношение по каждому слою приведено на диаграммах (рис. 2), анализ которых позволяет сделать ряд заключений.
1. За время накопления плейстоценовых слоев Денисовой пещеры видовой состав млекопитающих изменялся незначительно, что отражает, по-видимому, единый механизм формирования танатоценоза пещеры и неизменный состав населения мелких млекопитающих в долине Ануя в течение накопления изученной толщи. Это свидетельствует о единстве крупного палеогеографического этапа развития природной среды за период формирования плейстоценовых осадков пещеры.
