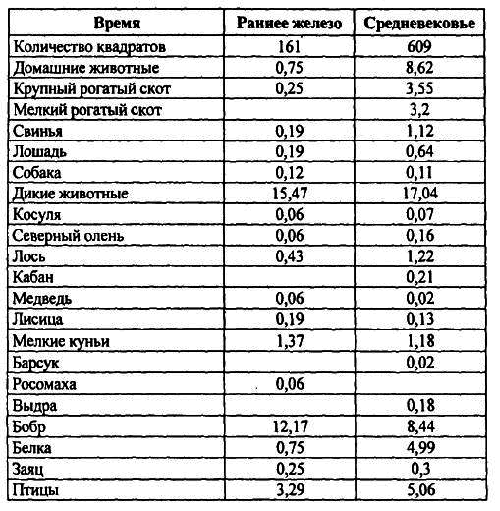
Сравнение изменения концентрации костей в раннежелезный век и средневековье показало (табл. 7), что количество остатков домашних животных в средневековье, по сравнению с раннежелезным векрмг увеличилось более чем в 10 раз, причем увеличение отмечается у всех домашних животных (за исключением собаки), хотя и в разной степени.
Таблица 7. Динамика концентрации костей (шт./м 3) в культурном слое Мининского археологического комплекса в раннежелезный век и средневековье.
Из основных промысловых видов трехкратное увеличение численности в средневековье отмечается у лося, что может быть связано с развитием подсечного земледелия и широким распространением вторичных лесов. Интересно отметить, что сходный порядок увеличения численности лося для этого периода получил и Н. К. Верещагин [ Верещагин, Русаков1979] для Прибалтики. Некоторое уменьшение численности отмечается у таких пушных зверей как бобр, лисица, мелкие куньи. Почти 7-кратное увеличение концентрации костей белки вряд ли связано со столь значительным изменением ее численности. Вероятно, это определялось большим ее преследованием из-за повышенного интереса к беличьим шкуркам как эквивалента денег, экспортного товара и т. п.
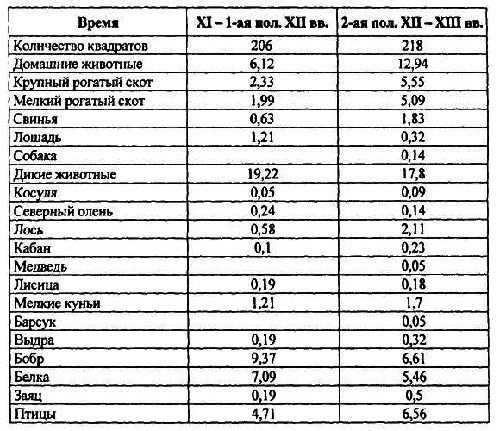
Таблица 8. Динамика концентрации костей (шт./м 3) в культурном слое Мининского археологического комплекса в средневековье.
Вычисление концентрации костей в различные периоды средневековья показало (табл. 8), что величина стада [8]домашних животных увеличилась во 2-ой половине XII–XIII вв., по сравнению с предыдущим периодом, почти в два раза. Двух- трехкратное увеличение численности отмечается у крупного и мелкого рогатого скота и свиньи. В тоже время отмечается резкое уменьшение роли лошади в питании населения. Аналогичные результаты получил и В. И. Цалкин [1956], который видит причину этого в изменении организации сельского хозяйства, т. е. перехода от подсечного земледелия к пашенному и использованию лошади преимущественно в качестве тягловой силы.
Из основных промысловых видов отмечается почти 4-кратное увеличение численности лося, что, как уже отмечалось, могло быть связано с увеличением площади вторичных лесов. Небольшое увеличение численности отмечается у мелких куньих, выдры, зайца. Незначительное уменьшение численности во 2-ой половине XII–XIII вв. было у бобра и белки.
Заключение
Чтобы сделать материалы современных археозоологов пригодными для экологических исследований следует охотничью добычу древних людей выражать скоростью накопления ее костей (СНК) на единице площади горизонтов культурного слоя, изучаемого поселения СНК = Nкв/tS. Для этого необходимо располагать их радиоуглеродными или точными археологическими датировками.
Помня, что люди добывали не кости, а животных вместе с их скелетом, важно по парным костям, обнаруженным в культурном слое, установить возможное число добытых особей и количество костей, потерявшихся до захоронения.
Относительную численность на охотничьем участке добывавшихся видов можно рассчитать по предложенной Чэпменом [ Chapman1974] модификацией метода Делюри \DeLury1947], связавшего среднюю численность промышляемого вида с его добычей и промысловыми усилиями, затраченными на ее получение — CPUE = Ct/flq. Из членов этого уравнения только ц — промысловый коэффициент или добычливость способа охоты, не оставляет следов в культурном слое и надежных способов оценки не имеет. Определяя численность промышлявшихся видов отношением Ct/flq и не принимая во внимание значение добычливости способов охоты, мы уравниваем влияние промысла на численность разных видов и не можем оценить влияние охоты на изменение их обилия. В таком виде CPUE отражает только влияние на этот процесс природных факторов. Чтобы исправить положение необходимо найти способы оценки промыслового коэффициента разных охот.
В районах развитого скотоводства и земледелия определение СРЦЕ значительно осложняется распределением основных промысловых усилий между этими основными отраслями хозяйства.
Благодарности
Мы искренне благодарим М. М. Бронштейна и К. А. Днепровского за предоставленный материал из древнеэскимосских поселений Чукотки и Н. А. Макарова за материал из Мининского памятника и его обсуждение. Также мы глубоко признательны Е. Е. Антипиной за ценные замечания и обсуждение рукописи.
Работа выполнена при финансовой поддержке РФФИ № 00-04-48149, № 03-04-49323 и Программы «Научные основы сохранения биоразнообразия России».
Литература
Арутюнов С. А., Сергеев ДА.Проблемы этнической истории Берингоморья (Эквенский могильник). М.: Наука, 1975.
Бекени Ш.Новый метод вычисления количества особей животных в остеологическом материале из археологических местонахождений // Бюл. МОИП. Отдел биологии. Т. 74. Вып. 6.1969. С. 69–71.
БронштейнММ Древнеэскимосское искусство как историко-этнографический источник: Автореф. дис… канд. ист. наук. М., 1990.
Верещагин Н. К., Русаков О. С.Копытные Северо-Запада СССР (история, образ жизни и хозяйственное использование). J1.: Наука, 1979.
Динесман Л. Г., Киселева Н. К., Савинецкий А. Б., Хасанов Б. Ф.Вековая динамика прибрежных экосистем северо-востока Чукотки, М.: Аргус, 1996.
Коли Г.Анализ популяций позвоночных. М.: Мир, 1979.
Крупник ИИАрктическая этноэкология. М.: Наука, 1989.
Ляпунова Р. Г.Алеуты и их экосистема // Экология американских индейцев и эскимосов. М.: Наука, 1988. С. 84–93.
Макаров Н. А.Шесть лет раскопок Мининского археологического комплекса // Кубенское озеро: взгляд сквозь тысячелетия. Вологда, 2001. С. 3–6.
Макаров НА., Захаров С. Д., Зайцева НЕ.Сельские поселения на Кубенском озере в XII–XIII вв. — от расцвета к запустению // Русь в XIII веке: континуитет или разрыв традиций? М, 2000. С. 65–72.
Паавер К. Л.Формирование териофауны и изменчивость млекопитающих Прибалтики в голоцене. Тарту, 1965.
Паавер К. Л.Методы оценки обилия субфоссильных костей ИЧастные методы изучения истории современных экосистем. М.: Наука, 1979. С. 212–222.
Рикер У. Е.Методы оценки и интерпретации биологических показателей популяций рыб. М.: Пищевая промышленность, 1979.
Савинецкий А. Б.Вековая динамика населения млекопитающих и птиц побережья и островов Берингова моря в последние тысячелетия. Автореф. дисс… докг. биол. наук. М: ИПЭЭ РАН, 2000.
Цалкин В. И.Материалы для истории скотоводства и охоты в древней Руси // Материалы и исследования по археологии СССР. № 51.1956.
Цалкин В. И.Древнейшие домашние животные Восточной Европы. М.: Наука, 1970.
Acsadi G., Nemeskeri J.History of human life span and mortality. Budapest: Academical Kiedo, 1970.
Allen KRSome methods for estimating exploited population // J. Fish. Res. Board Can. Vol. 23. N 10. 1966. P. 1553–1574.
Allen KR, Kirkwood G. P.Man impacts on marine mammals // Fish population dynamics. Chichester, etc. 1988. P. 151–269.
Bockstoce JR., Botkin D. B.The historical status and reduction of the Western Arctic bowhead whale population by the pelagic whaling industry // Rep. Int. Whal. Comm. Special issue 5.1983.P. 107–142.
Chapman D. G.Estimation of population size and sustainable yield of sei whales in the Antarctic // Rep. Intern. Whaling Commiss. V. 24. 1974. P. 82–90.
Chatters J. C.The faunal analysis of sites MK2 and MK4 Nunivak, Alaska. Report from Univ. of Washington. 1972. R 1-21.