Объектом нашего изучения были кости из культурных слоев поселений древних эскимосов и алеутов, расположенных на побережье и о-вах Берингова моря, а также культурного слоя из Мининского археологического комплекса (Вологодская обл.), формировавшегося с мезолита до средневековья [ Макаров2001].
1. Древние поселения на побережье и островах Берингова моря
Азиатские эскимосы занимают узкую полосу приморских тундр, сформировавшихся под влиянием местной циркуляции морского воздуха. Алеуты живут на о-вах Алеутской гряды. И древние эскимосы, и древние алеугы жили в долговременных полуземлянках, по руинам которых сейчас легко обнаружить места былых поселений этих народов.
Каждое из древних поселений эскимосов и алеутов имело свой охотничий участок и представляло собой относительно автономную хозяйственную, социальную и репродуктивную ячейку [ Ляпунова1988; Крупник1989].
Следует подчеркнуть, что эскимосы и алеуты были охотниками-собира-телями, никогда не занимавшимися скотоводством и земледелием. Здесь изучение влияния охотничьего хозяйства на природные экосистемы представляет особый интерес. В других районах России его следы стерты позднее возникшими формами использования природных ресурсов — скотоводством, земледелием и другими видами хозяйственной деятельности.
Как показали серийные радиоуглеродные датировки [ Динесман и др.1996; Dinesman et al.1999; Савинецкий2000] поселения алеутов и азиатских эскимосов существовали в течение 2–3 тысяч лет.
1.1. Древние поселения эскимосов и алеутов и особенности культурных слоев этих памятников
На участке, занятом поселением, люди быстро вытаптывали естественный травяной и моховой покров, разрушали структуру верхнего горизонта почвы и постоянно замусоривали его бытовыми отходами. Кучи их возникали у входов каждой полуземлянки, откуда постепенно растаскивались по всему участку. Сам участок периодически заносился супесями, песком или суглинками и промачивался атмосферными осадками. Это приводило к развитию локального почвообразовательного процесса. Обычно культурные слои богаты фосфором и имеют темную окраску. Это связано не только с высоким содержанием гумуса, но и с наличием мельчайших частиц угля, вымытых из кухонных отбросов.
Мощность культурного слоя определяется по глубине захоронения в нем костей животных и артефактов. В древних поселениях эскимосов и алеутов она нередко достигает 1–1,5 м.
Если поселение покидается людьми, культурный слой снова зарастает растительным покровом, в котором значительную роль играют полынь и сорные виды, устойчивые к вытаптыванию и особенностям химического состава культурного слоя. За их счет даже поселения, покинутые людьми 200–600 лет назад, теперь еще выделяются на окружающем фоне ярко зелеными цветом.
Если покинутые поселения вновь заселяются людьми, то их жизнедеятельность приводит к возникновению нового культурного слоя. От первого погребенного культурного слоя он отделяется горизонтом суглинка, супеси или песка, накопившихся во время перерыва использования поселения людьми. Чтобы убедиться в отсутствии погребенных культурных слоев, разведочные шурфы углубляются до почвообразующей породы.
1.2. Методика сбора и количественного учета костей в культурном слое
Для сбора и учета костей, сохранившихся в культурном слое изученных нами памятников, его горизонты раздельно просеивали, а если позволяла обстановка, то промывали на ситах водой.
При отборе проб из горизонтов для просеивания или промывки необходимо тщательно следить за положением их границ Это позволяет избежать перемешивания геологически разновозрастного остеологического материала. Чтобы облегчить себе эту работу, раскопку культурного слоя следует начинать двумя взаимно перпендикулярными шурфами, на зачищенные стенки которых наносят границы выделяемых горизонтов. Желательно, чтобы хотя бы часть из них совпадала с границами почвенных горизонтов. Счет погребенных культурных слоев и их горизонтов ведется от нижней части заложенных шурфов вверх. Глубина их залегания измеряется от поверхности современной почвы или от условного нулевого уровня.
После отбора костей и их определения выбирается тот количественный показатель, который больше подходит к соответствующему типу археологического памятника, а также в зависимости от целей исследования.
Наиболее часто археозоологи учитывают общее число найденных костных остатков вида (ОКО) и минимальное число особей (МЧО), которым они могли принадлежать [ Бекени1969; Цалкин1970; Паавер1979; Klein, Cruz-Uribe1984; и др.].
Чаще всего минимальное число особей подсчитывается по наиболее многочисленному элементу скелета. Желательно при этом учитывать индивидуальную и возрастную изменчивость элементов скелета [ Бекени1969]. В остеологических выборках соотношение правых и левых элементов какой-либо кости, как и в скелете позвоночных, должно быть близко к 1.
Таблица 1. Парные кости (шт.*) птиц из культурного слоя Дежневского древнеэскимосского поселения.
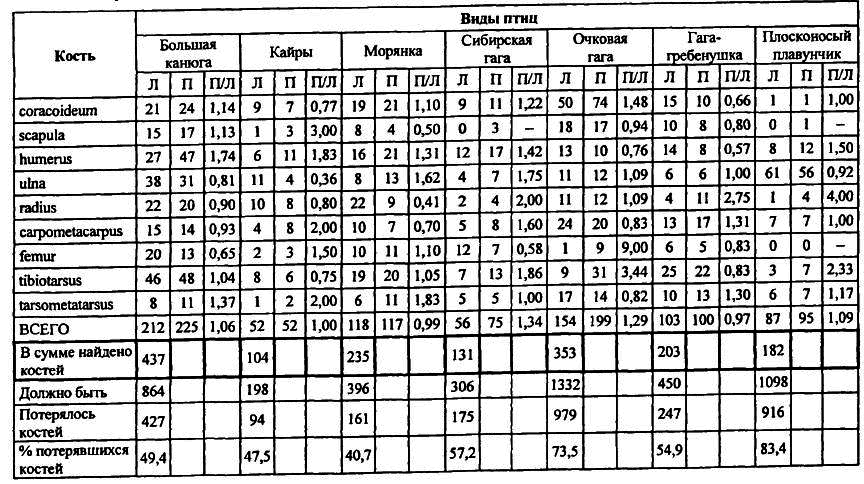
В выборках костей из культурного слоя Дежневского древнеэскимосского поселения Чукотки (табл. 1) это нередко имеет место. Помня, что охотники добывают не кости, а животных с их целыми скелетами, приходится признать, что отклонение в выборках численного соотношения правых и левых костей от нормальной величины связано только с потерей части добытого материала. Она могла произойти при разделке туш животных и их дальнейшей подготовке к использованию. Оценить потери можно стандартизировав остеологическую выборку по анатомическим нормам скелета животных [ Динесман и др.1996; Dinesmanеt а1.1999]. Для этого достаточно подсчитать в ней общее количество всех крупных разноименных костей поясов конечностей вида (Mкв) и по наиболее многочисленной из них определить обычным способом минимальное число особей (МЧО). Количество (А) разноименных парных костей поясов конечностей, приходящееся на одну только что добытую особь вида, зависит от набора элементов скелета, выбранных для таксономической диагностики остеологического материала. Оно нам хорошо известно и всегда больше аналогичного показателя Мкв/МЧО, подсчитанного для выборки. Разница этих величин определяется потерей части костей добычи (Мук) еще до захоронения в культурном слое. Очевидно, исходное количество парных костей зафиксированного выборкой минимального числа особей (Мисх) равняется сумме общего числа ее костей и потерявшихся костей, т. е. Мисх = Мкв + Кук. Эго позволяет считать, что (Мкв + Мук)/МЧО = А, или Кук = А × МЧО — Мкв. В качестве примера можно привести следующий. Наиболее многочисленным найденным элементом скелета большой конюги был тибиотарзус (М = 48 правых костей) (табл. 1). Благодаря двусторонней симметрии скелета, такое же число левых тибиотарзусов должны были иметь и все другие большие конюги, добытые дежневскими эскимосами. Таким же должно было быть у них количество правых и левых элементов каждой крупной парной кости конечностей. Очевидно, для определения суммы добытых костей большой конюги нужно 48 × 2 (число тибиотарзусов у одной особи) × 9 (число наименований парных костей, использованных для таксономической диагностики выборок) — 48 × 2 × 9 = 864. Таким образом, добытые большие конюги имели 864 парные кости девяти наименований. Вычтя из этого числа сумму всех найденных правых и левых парных костей большой конюги (437) (табл. 1), получим 864–437 = 427 или 49 % костей этого вида потерялось при обработке добычи.
Выполнение аналогичных вычислений для других видов промысловых птиц добывавшихся эскимосами, показало (табл. 1), что еще до захоронения в культурном слое теряется от 41 % до 84 % костей. Такие большие потери костей не должны удивлять. У древних эскимосов кости добычи не только попадали в культурный слой вместе с кухонными отходами, но и использовались для изготовления оружия и бытовых предметов, шли на сооружение каркасов землянок и мясных ям, сжигались как топливо, растаскивались собаками.