Кроме хорошо известных современных зональных комплексов, сложившихся в результате развития фаун в голоцене, в позднем плейстоцене существовали фауны, в отношении зональной принадлежности которых единого мнения нет. Разные точки зрения на этот предмет были рассмотрены нами недавно в специальной работе [ Смирнов2001], а здесь больше внимания будет уделено преобразованиям сообществ позднего валдая в голоценовые и современные.
Имеется два подхода к пониманию сути зонального распределения биоты в позднем валдае: 1) оно понимается как мозаика современных биотопов из разных зон, в результате чего формируется смешение черт современных зональных комплексов (тундростепь); 2) принимается своеобразие зоны, которую невозможно свести к «гибриду» зон, существующих ныне.
Назрело введение нового названия для плейстоценового зонального комплекса, который не имеет прямых современных аналогов, дисгармоничен с точки зрения современных зональных сообществ и обладает рядом черт как степного, так и тундрового комплексов, знаковым видом среди крупных млекопитающих в нем является мамонт, среди мелких — копытный лемминг. Этот комплекс занимал большую часть поздневалдайской гиперзоны, то есть северной (бореальной) части Евразии, что делает привлекательным обозначить его как «гиперборейный» [ Смирнов2001].
Рассмотрим зональное распределение фаун млекопитающих позднего валдая на примере востока Урала и положение гиперборейной зоны на этой территории относительно других зон. Самым северным был поздневалдайский арктическийкомплекс, он отличался бедным составом и максимально упрощенной структурой. Среди мелких млекопитающих резко преобладал копытный лемминг Dicrostonyx guilielmi. Например, в местонахождении Пымва-Шор (западное Предуралье, широта 67° с.ш.), среди нескольких тысяч зубов из разных горизонтов, доля копытного лемминга составила 90–95 %, сибирского 5-10 %, а узкочерепной полевки — единицы процентов. Основные свойства этого комплекса были обусловлены не только низкими температурами, но и большей сухостью климата. Среди крупных млекопитающих заметно преобладали северный олень, овцебык и лошадь; из них первые два вида, безусловно, можно отнести к зональным. Мамонт и бизон также встречались, но их, как и лошадь, скорее следует отнести к группе экстразональных, так как их массовые находки и, вероятно, основной ареал находятся южнее. Комплекс был распространен к югу от края ледника и побережья океана на севере, его южная граница проходила между 63 и 65°с.ш.
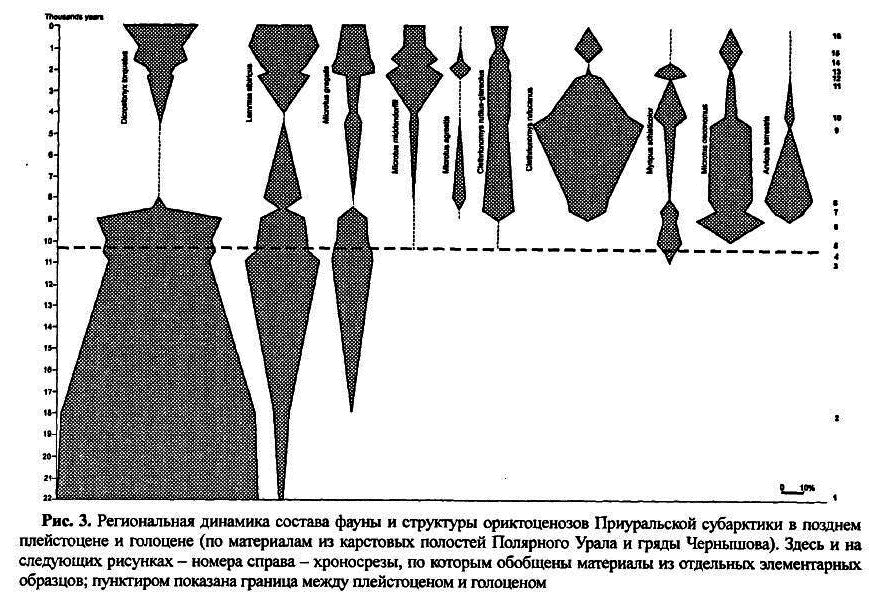
На рисунке 3 изображена динамика состава и структуры населения грызунов в позднем валдае и голоцене, реконструированная на основе исследования 16 элементарных образцов из карстовых полостей Полярного Урала и гряды Чернышова [ Смирнов, Андреичева и др.1999; Smirnov, Golovachev А999; Головачев1998; 1999; 2000]. Всего при построении этой диаграммы было изучено более 10 тысяч щечных зубов грызунов. На диаграмме видно, что для этого региона единственное эволюционное преобразование фауны произошло примерно через тысячу лет после плейстоцен-голоценового рубежа и было очень резким. Ему предшествовали смены внутри арктического комплекса, исторического масштаба, — от абсолютного доминирования копытного лемминга в позднем валдае, с нарастанием долей сибирского лемминга и узкочерепной полевки к позднеледниковью. Уже примерно 8 тысяч лет назад полностью сформировался субарктический комплекс, в котором видовое богатство выросло в три раза. Доминанты сменились до такой степени, что в середине голоцена таковым стала красносерая полевка при почти полном исчезновении леммингов из состава комплекса. Позднее, в течение последних 4 тысяч лет, снова стали расти доли зональных тундровых видов: леммингов, узкочерепной полевки и полевки Миддендорфа, но едва ли следует считать, что произошло второе преобразование эволюционного уровня. Хотя произошла новая смена доминантов, которыми снова стали упомянутые зональные тундровые виды, но лесные виды в качестве экстразональных в составе фауны сохранялись, и комплекс в целом на протяжении всего голоцена имел лесотундровый облик.
Гиперборейный комплекс млекопитающихбыл распространен там, где сейчас находится вся зона тайги, зона смешанных лесов и лесостепь. Главным свойством этого комплекса было большое видовое разнообразие, достигающее максимума в центральной части зоны. К зональным видам гиперборейного комплекса можно отнести мамонта, шерстистого носорога, пещерного медведя, лошадь. Большинство из них вымерли вместе с исчезновением гиперборейной зоны и распадом комплекса. Среди мелких млекопитающих едва ли можно назвать хотя бы один вид, свойственный только этой зоне, — то есть, все они были полизональными. Совместное присутствие в этом комплексе видов, ныне обитающих в тундре, степи и полупустыне, безусловно, является следствием своеобразия климата. Определяющее его свойство — большая сухость при низких зимних и умеренных летних температурах. Комплекс не был однородным; выделяются как минимум три подзональных варианта. Комплекс северной гиперборейной подзоныбыл через переходную полосу связан на севере с поздневалдайской арктической зоной. Самые северные местонахождения, которые можно отнести к этому варианту гиперборейного комплекса, известны с верховьев Печоры (Медвежья и Студеная пещеры, 62° с.ш.), но, вероятно, они могут быть обнаружены и севернее, на 63–64° с.ш. Южная граница этой подзоны проходит примерно по 60° с.ш. Ядро населения мелких млекопитающих в этой подзоне составляли позднеплейстоценовый копытный и сибирский лемминги, хотя первый заметно преобладал; в локальных фаунах сумма их остатков превышала 60 %. Обилие копытного лемминга сближает описываемый комплекс с тундровым. Однако он отличается от последнего большим видовым богатством, присутствием степной пищухи, большим участием узкочерепной полевки и экономки, а также редкими находками степной пеструшки и полевок рода Clethrionomys.Среди крупных млекопитающих в этой подзоне наиболее массовыми были северный олень, овцебык и лошадь, при заметном количестве пещерного медведя, мамонта и бизона. Переход к голоцену на севере распространения гиперборейного комплекса был не столь резким, как в Арктике, и пока еще не достаточно материалов для того, чтобы охарактеризовать его с необходимой подробностью.
Комплекс типичной гиперборейной подзоныизвестен по многим местонахождениям Северного и Среднего Урала, от 60° до 56° с.ш. На рисунке 4 показаны доли основных видов в его составе на примере региональной фауны западного склона Среднего Урала. Для этого комплекса характерно высокое видовое разнообразие, при доминировании узкочерепной полевки (30–40 %). Вместе с копытным леммингом эти два вида составляли основу подзонального комплекса. Степная пеструшка, экономка и сибирский лемминг в разных локальных фаунах занимают также весьма заметное место. В локальных фаунах по всей подзоне устойчиво встречаются темная и водяная полевки, красная и красно-серая полевки, серый хомячок, степная пищуха. На контакте с южной подзоной известны единичные находки тушканчиков и желтой пеструшки; это явно экстразональные элементы, проникающие с юга. Среди крупных млекопитающих в этой подзоне встречались шерстистый носорог, пещерный медведь, мамонт, лошадь, северный олень, бизон и реже овцебык.
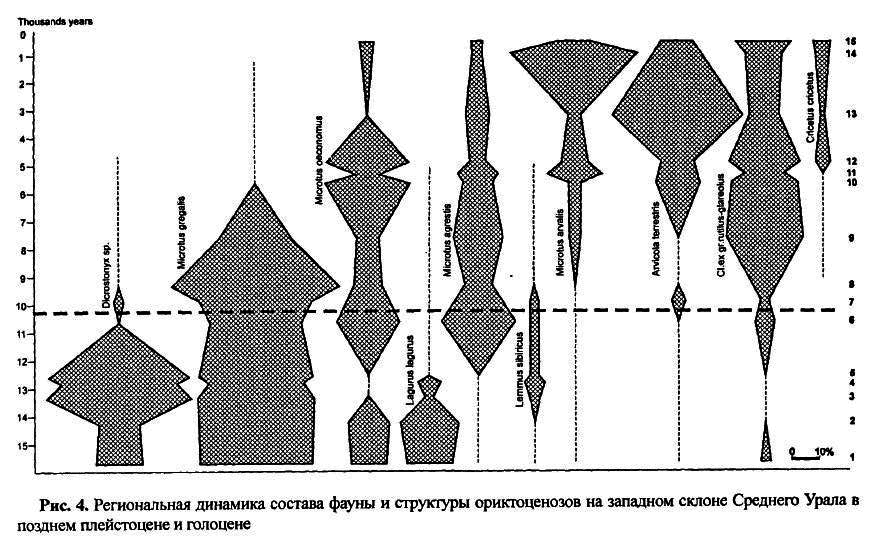
Региональная динамика этого комплекса, которая к нашему времени привела к формированию современного комплекса мелких млекопитающих, характерных для южной тайги с преобладанием лесных полевок и заметной долей полевок надвида обыкновенной полевки. Эта динамика прослежена на основании обобщения данных по 15 хроносрезам (рис. 4). Для характеристики были привлечены данные по 40 элементарным образцам, с общим числом щечных зубов мелких млекопитающих более 100 тысяч [ Смирнов1992; 1993; 1995; Фадеева, Смирнов и др.2000].