Конкретный пример такого усреднения удалось получить благодаря тому, что в заповеднике орнитологами по содержимому погадок велось изучение питания филина; есть сведения за 30–40 годы ХХ-го века и 86–93 гг. [ Астафьев, Нейфельд1999]. Там же (на берегу р. Печоры в районе устья р. Шежим) И. Б. Головачевым и Н. О. Садыковой по нашей просьбе были собраны субфоссильные остатки грызунов из мест многолетней концентрации погадок филина (за передачу этих сборов для обработки автор выражает коллегам искреннюю благодарность). Эти костные остатки представляют собой элементарный образец из отложений средней мощностью 10 см. Точно определить временной отрезок накопления образца невозможно, но, судя по степени сохранности костей, глубине и характеру их залегания, а также наличию в нем остатков ондатры, время формирования полученного материала, вероятно, несколько превышало последние сто лет. Необходимо отметить, что погадки накапливались не равномерно во все годы наблюдения, а лишь в годы успешного гнездования филина, соответствующие периодам высокой численности их кормовых объектов. 30-40-е годы пришлись на высокую численность белки и зайца-беляка, которые и преобладали (среди млекопитающих) в пробах питания. В конце ХХ-го века самым успешным для гнездования был 1989 год, когда белки было мало (доля в составе добычи 2,7 %), но зато наблюдалась высокая численность лесного лемминга и ряда видов полевок. Многолетние данные отловов грызунов разными способами показали, что в целом в лесных биотопах района среди грызунов преобладают красная и рыжая полевки, при периодических вспышках численности лесного лемминга [Закономерности… 2000]. В остатках питания филина доминировали лесной лемминг, темная, водяная и красно-серая полевки (доля каждого вида среди млекопитающих более 10 %).
В субфоссильном образце доминировали остатки белки (29 %), примерно в равных долях (по 8-11 %) были представлены остатки водяной, темной, полевки экономки, лесного лемминга. Полевки трех видов рода Clethrionomysв сумме составили 13 %.
Таким образом, в элементарном образце причудливо отразилась картина динамики населения, через механизм накопления остатков не во все годы, а преимущественно в годы высокой численности животных, способных обеспечить успешное гнездование хищника. Поскольку за время накопления элементарного образца случаются периоды вспышек численности разных видов, происходит усреднение облика локального населения в орикгоценозе.
Локальная фауна и локальный ориктоценоз
Следующий уровень в алеалогическом исследовании, после описания элементарных образцов, элементарных фаун и ориктоценозов, — выделение и описание более общих единиц Это локальные фауны и соответствующие им локальные ориктоценозы. В основе процедуры выделения локальной фауны — обобщение нескольких элементарных образцов, которые могут происходить из одного разреза или разных, но синхронных, расположенных в одной местности; сходное определение предложили ранее Р. Грэхем и Х. Семкен [ Graham, Semken1987]. В пределах одной локальной фауны может наблюдаться динамика определенного уровня. Необходимо специально рассмотреть, какие вариации элементарных фаун следует считать происходящими в пределах одной локальной фауны, и как определить границы перехода состояния населения территории на новый качественный уровень, чтобы фиксировать смену одной локальной фауны на другую.
Если локальная фауна за столетия и тысячелетия претерпевает лишь обратимые флуктуации относительной численности отдельных видов, не создающие устойчивых трендов, приводящих к смене доминантов, и при этом сохраняется ее видовой состав, то такую динамику следует относить к актуальному масштабу. В отличие от процессов более высокого уровня, такую динамику далее будем называть изменениями. Пример изменений примерно за тысячелетний промежуток времени, прослеженных в пределах одного слоя на выборках очень большого объема, можно видеть в таблице 7.
Когда динамика состава фауны и структуры приобретает определенную устойчивую направленность, то можно говорить о смене в историческом масштабе. Ее результат — такой сдвиг относительной численности видов, который приводит к смене характерных черт конкретной локальной фауны, когда ее уже нельзя считать прежней и необходимо признать сменуодной локальной фауны (и соответствующего ей ориктоценоза) на другую. Как во всяком непрерывном процессе, провести линейную границу между той и другой фауной бывает трудно. Конечную стадию процесса удобно фиксировать по смене доминирующих по относительной численности видов. К сменам в историческом масштабе предлагается относить не все случаи динамики со сменой доминантов, а только такие, когда доминанты меняются в пределах одного зонального типа, и кардинального изменения видового состава еще не наблюдается. Можно выделить два уровня смен локальных фаун — начальный и продвинутый.
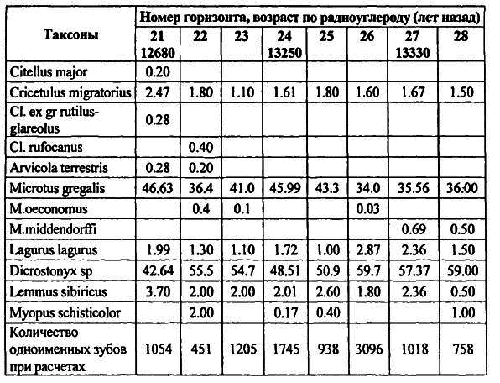
Таблица 7. Доли видов и число щечных зубов грызунов в условных горизонтах 21–28 слоя 10 грота Расик [ Фадеева, Смирнов и др.2000] (%)
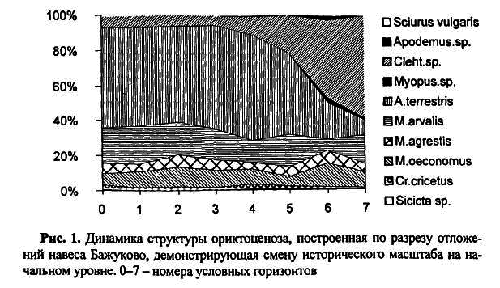
В качестве примера первого можно привести динамику состава фауны и структуры населения грызунов в позднем голоцене, прослеженные на материалах навеса Бажуково на Среднем Урале. Отложения в этом навесе вскрывались условными горизонтами по 5 см. На рис. 1 видно, что на изученном отрезке времени видовой состав остается постоянным. Наблюдается устойчивый тренд сокращения относительной численности полевок рода Clethrionomysи возрастание доли водяной полевки. Но поскольку общий южнотаежный зональный облик фауны сохраняется, а меняется лишь соотношение относительной численности зональных таежных видов и азональных обитателей лугов, то можно сделать вывод о том, что наблюдаемая динамика относится к начальному этапу исторических смен локальной фауны.
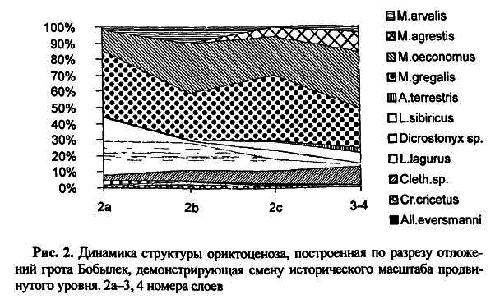
Другой пример демонстрирует продвинутый уровень исторической смены локальной фауны. На рис. 2 показана динамика состава фауны и структуры населения мелких млекопитающих, прослеженная по 5 слоям поздневалдайских отложений в гроте Бобылек [ Смирнов1993]. Верхний слой 2а имеет радиоуглеродный возраст 14 200 лет, а дата для слоя 3 — более 33 тыс. лет. На всем представленном отрезке разреза локальная фауна соответствует одному зональному типу. Это гиперборейная фауна, в которой копытные и сибирские лемминги соседствуют со степной пеструшкой и узкочерепной полевкой. Тем не менее, от слою к слою заметны сдвиги не только долей остатков видов разных групп, изменяется и видовой состав, в верхней части отложений ее характеристики ближе к типичной гиперборейной подзоне, а в нижних слоях нарастает доля лесных и луговых элементов, уменьшается роль степной пеструшки; узкочерепная полевка теряет доминирующее положение, исчезает ряд видов, типичных для аридных местообитаний, и наоборот, появляются мыши, белка и обыкновенный хомяк, которых не было в верхних слоях.
Региональный комплекс — это совокупность одновременных локальных фаун, характеризующая более широкий географический район.
Процедура перехода от изучения локальных фаун к региональному комплексу зависит от степени насыщенности материалом изучаемого отрезка времени и количества исследованных местонахождений на исследуемой территории: она будет описана ниже.
Позднеплейстоцен-голоценовая динамика региональных и зональных фаунистических комплексов на Урале — восточной окраине Европы
Кроме рассмотренных смен и изменений, локальные и региональные фауны мелких млекопитающих в позднем плейстоцене и голоцене проходили стадии динамики большего масштаба — эволюционного. Они связываются с преобразованием в другой зональный тип. Критериями такого преобразования могут служить смена видов-доминантов (если они относятся к разным зональным группам) и существенное изменение видового состава.