Всю прослеженную динамику можно разделить на два крупных этапа. Переход от одного к другому, как ни странно, проходит не по рубежу плейстоцена и голоцена, а приурочен к началу голоценового оптимума (около 7,5 тыс. лет). Именно здесь можно провести эволюционный рубеж в преобразовании регионального комплекса мелких млекопитающих. В течение всего первого этапа видом доминантом в ориктоценозах была узкочерепная полевка, уступившая на втором этапе роль доминанта лесным полевкам. Внутри первого этапа можно выделить три фазы развития, которым следует придать статус исторических смен. Эти фазы выделяются по смене субдоминантов. На первой фазе (16–14,5 тыс. лет) второе и третье место по относительной численности занимали копытный лемминг и степная пеструшка. На второй (14,5-11 тыс. лет) роль степной пеструшки в составе орикгоценоза резко сократилась, а копытный лемминг в середине этой фазы вышел на место содоминанта с узкочерепной полевкой. Далее его относительная численность резко сокращалась, и с плейстоцен-голоценового рубежа до середины голоценового оптимума его присутствие в составе ориктоценозов было едва уловимо. Третья фаза может быть обозначена как фаза деградации гиперборейного комплекса. Она продолжалась примерно до 7,5 тыс. лет. В ее начале узкочерепная полевка была сверхдоминантом (доля ее остатков превосходила 60 %), но затем, примерно за 2 тысячи лет она сократилась до 20 %. Степная пеструшка, как и ряд других видов из состава гиперборейного комплекса (сибирский лемминг, серый хомячок, степная пищуха) еще встречались, но, как и копытный лемминг, перешли в категорию редких видов.
С переходом ко второму (лесному) этапу развития региональный комплекс претерпевал несколько исторических смен. Сначала в нем доминировали зональные таежные виды с небольшой примесью экстазональных (степных), а затем азональных (луговых). Всплеск относительной численности Microtus arvalisможно трактовать как следствие развития агроландшафтов.
Комплекс южной гнперборейной подзоныобнаружен во многих пещерах горной части Южного Урала [Историческая экология… 1990]. Его северная граница совпадает с южным краем типичной подзоны, а южная — требует уточнения. Наиболее вероятно, она проходит на Урале по 52° с.ш. Для этого подзонального варианта гиперборейного комплекса на региональном уровне характерно резкое доминирование узкочерепной полевки (до 70 %), обилие экономки и степной пеструшки, при постоянном присутствии серого хомячка и копытного лемминга (до 10 %) и реже — сибирского лемминга. В составе регионального варианта комплекса отмечены также хомячок Эверсманна, два вида тушканчиков и два вида сусликов, степная пищуха, а на восточном склоне слепушонка. С его распадом в нижних поясах гор Южного Урала сформировался к современности южнотаежный комплекс, с примесью элементов европейских широколиственных лесов (желтогорлая мышь и садовая соня).
Фауны лесостепного, степного или полупустынногокомплексов в позднем плейстоцене не имели в своем составе ни копытных, ни сибирских леммингов, а другие виды мелких млекопитающих — почти все общие с южным вариантом гиперборейного комплекса. Они известны в Предуралье, Зауралье и Западной Сибири [ Зажигин1980; Малеева, Стефановский1988; Кузьмина, Смирнов и др.2001; Кузьмина2002]. В полевом сезоне 2002 года нами совместно с Е. А. Кузьминой и А. И. Улитко в Южном Зауралье, на междуречье рек Урал и Большой Кизил, были раскопаны отложения поздневалдайского и голоценового возраста в пещере Сыртинская с массовым остеологическим материалом и по крупным, и по мелким млекопитающим. На рисунке 5 приведены предварительные данные о динамике сообществ мелких млекопитающих Южного Зауралья в пределах современной степной зоны, полученные на базе обобщения сборов из 14 элементарных образцов, 4 местонахождений. Общее количество исследованных щечных зубов около 15 тыс. Первый хроносрез охарактеризован по материалам из условного горизонта 15 пещеры Сыртинская.
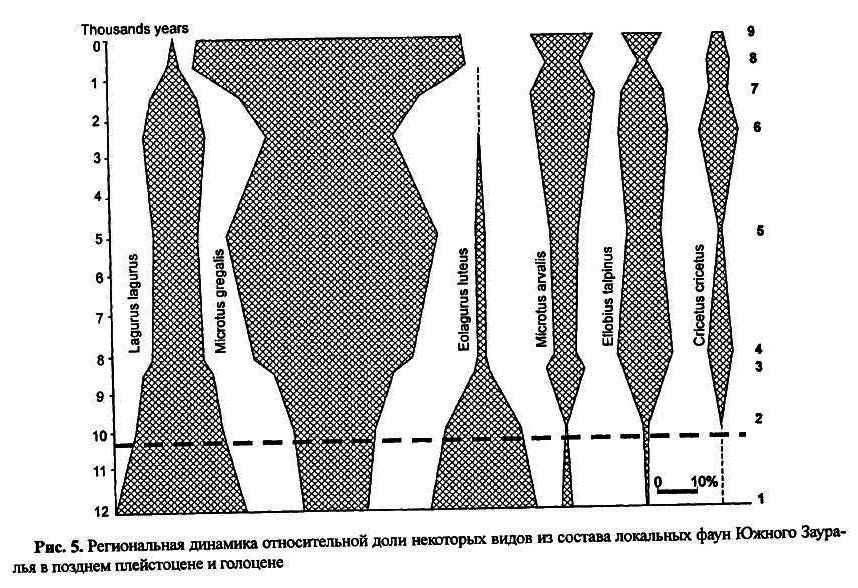
По составу доминатнов эту фауну (рис. 5, 1) можно назвать лагурусно-эолагурусной, а узкочерепная полевка занимает в ней по относительной численности третье место; ее следует отнести к типу фаун сухих степей и полупустынь. На плейстоцен голоценовом рубеже кардинальной трансформации фауны не наблюдается. В интервале от 10 до 8 тыс. лет происходит заметное сокращение доли остатков желтой пеструшки, небольшое уменьшение доли степной пеструшки и рост относительной численности слепушонки, обыкновенного хомяка, обыкновенной полевки. Узкочерепная полевка увеличивала относительную численность вплоть до 5 тыс. лет. На протяжении почти всего голоцена региональную фауну можно называть грегалисно-лагурусной, и только в историческое время она трансформируется в грегалисно-арвалисную. Весь комплекс имеющихся материалов позволяет предположить, что в южно-зауральском регионе процесс преобразования фауны мелких млекопитающих позднего валдая растянулся почти на весь голоцен и только к нашему времени достиг эволюционного уровня. Дальнейшие исследования в этом регионе должны уточнить масштабы динамики на отдельных хроносрезах.
* * *
В позднеплейстоцен-голоценовой динамике состава фаун и структуры населения грызунов на разных широтах Урала прослеживаются процессы трех масштабов актуального, исторического и эволюционного.
Преобразования эволюционного масштаба сильно варьируют по хронологической протяженности; они могут проявляться как резкий скачок (от одной-двух тысяч лет) или быть растянуты (более десяти тысяч лет).
Смены исторического масштаба, условно подразделяемые на начальный и продвинутый уровни, по хронологической протяженности также существенно неоднородны. Их отдельные фазы с сохранением тренда определенного направления могут длиться в течение нескольких тысяч лет, но могут быть и существенно короче — всего несколько столетий.
Изменения актуального масштаба постоянно «вложены» в динамику большего масштаба, и разграничить их можно только по результатам и в исторической перспективе.
Благодарности
В подготовке данной работы принимали участие многие коллеги и ученики автора, и всем им автор приносит свою сердечную благодарность. Особенно хотелось бы выделить роль И. Б. Головачева, Е. А. Кузьминой, С. С. Трофимовой, Г. В. Быковой, А. И. Улитко и Н. О. Садыковой. Каждый из них по-своему помог автору на разных этапах сбора и подготовки материалов и оформлении рукописи. Работа выполнена при поддержке грантов: РФФИ 02-04-49181, и Российско-Нидерландского проекта по исследованию плейстоцен-голоценового перехода. Радиоуглеродные (АМ8) — датировки были получены благодаря поддержке гранта EAR-9018958 (by National Science Foundation).
Литература
Астафьев А. А., Нейфельд Н. Д.Род Bubo,филины // Фауна европейского Северо-Востока России. Птицы. Т.1.4.2. СПб., 1999.
Головачев И. Б.Голоценовые грызуны из грота «Зверобой» на Полярном Урале // Современные проблемы популяционной, исторической и прикладной экологии. Екатеринбург, 1998.
Головачев И. Б.Новые данные по динамике фауны грызунов Полярного Урала в голоцене (местонахождения Янгана-Пэ-3, Наун-Пэ) // Развитие идей академика С. С. Шварца в современной экологии. Екатеринбург, 1999.
Головачев И. Б.История фауны грызунов Приуральской Субарктики в голоцене // Биосфера и человечество. Екатеринбург, 2000.
Громов ИМ.Ископаемые верхнечетвертичные грызуны предгорий Крыма. М., 1961.
Завадский К. М., Колчинский Э. И.Эволюция эволюции. Л., 1977
Зажигин B. C.Грызуны позднего плиоцена и антропогена юга Западной Сибири. М.: Наука, 1980.
Закономерности полувековой динамики биоты девственной тайги Северного Предуралья / Бобрецов А. В., Бешкарев А. Б., Басов В. А., и др. Сыктывкар, 2000.