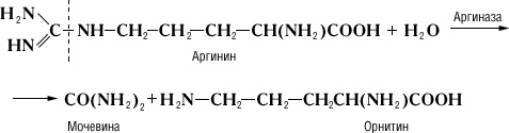Примером двухкомпонентного Ф. является пируватдекарбоксилаза , катализирующая расщепление пировиноградной кислоты на двуокись углерода и уксусный альдегид: CH3 COCOOH ® CH3 CHO + CO2 . Простетическая группа пируватдекарбоксилазы (тиаминнирофосфат) образована молекулой тиамина (витамина B1 ) и двумя остатками фосфорной кислоты. Простетические группы ряда важных окислительно-восстановительных Ф. – дегидрогеназ содержат производное амида никотиновой кислоты (ниацина), или же рибофлавина (витамина B2 ); в состав простетических группы т. н. пиридоксалевых ферментов , катализирующих перенос аминогрупп (–NH2 ) и декарбоксилирование и ряд др. превращений аминокислот, входит пиридоксальфосфат – производное витамина B6 ; активная группа Ф., катализирующих перенос остатков различных органических кислот (например, ацетила CH3 CO–), включает витамин пантотеновую кислоту . К двухкомпонентным Ф. относятся также важные окислительные Ф. – каталаза (катализирует реакцию разложения перекиси водорода на воду и кислород) и пероксидаза (окисляет перекисями различные соединения, например полифенолы с образованием соответствующего хинона и воды). Каталитическое действие этих Ф. может быть воспроизведено с помощью ионов трёхвалентного железа. Эти ионы обладают, однако, очень малой каталитической активностью, которая может быть усилена, если атом железа входит в состав гема . Хотя гем обладает уже значительным каталазным действием, его каталитическая активность всё же в несколько миллионов раз меньше активности каталазы, в которой гем в качестве простетической группы этого Ф. связан со специфическ им белком. Гем обладает также слабым пероксидазным действием, однако это действие проявляется в полной мере только после соединения гема со специфическим белком в целостный Ф. пероксидазу. Т. о., соединение простетической группы с белком приводит к резкому возрастанию её каталитической активности. Вместе с тем от природы белка зависит не только каталитическая активность, но и специфичность действия Ф. Прочность связи простетической группы и апофермента различна у разных Ф. У некоторых Ф., например у дегидрогеназ, катализирующих окисление различных субстратов путём отщепления водорода, эта связь является непрочной. Такие Ф. легко диссоциируют (например, при диализе ) и распадаются на простетическую группу и апофермент. Простетические группы, легко отделяющиеся от белковой части Ф., называются коферментами .
Многие Ф. содержат металлы, без которых Ф. не активен. Эти металлы называются кофакторам и. Так, пероксидаза и каталаза содержат железо, аскорбинатоксидаза, катализирующая окисление аскорбиновой кислоты, – медь, алкогольдегидрогеназа, окисляющая спирты в соответствующие альдегиды, – цинк.
Специфичность и механизм действия ферментов. Действие Ф., в отличие от неорганических катализаторов, строго специфично и зависит от строения субстрата, на который Ф. действует. Прекрасным примером такой зависимости служит катализируемая аргиназой реакция гидролитического расщепления аминокислоты аргинина на орнитин и мочевину:
Однако аргиназа не расщепляет метилового эфира аргинина:
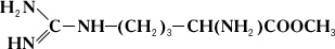
Дипептид, состоящий из остатков двух молекул аргинина, под действием аргиназы даёт лишь половину теоретического количества мочевины. Очевидно, что, хотя расщепление аргинина происходит в месте, весьма отдалённом от карбоксильной (COOH) группы (показано пунктиром), необходимым условием действия аргиназы является её соединение с карбоксильной группой аргинина. Поэтому замещение водорода в карбоксильной группе на метильный остаток или же связывание карбоксильной группы со второй молекулой аргинина оказывают резкое влияние на действие аргиназы. Примеры специфичности действия Ф. могут быть приведены при рассмотрении их стереохимической специфичности, т. е. действия Ф. на стереоизомеры (см. Изомерия ). Так, Ф., окисляющий природные L-аминокислоты, не действует на D-изомеры этих же аминокислот; Ф. дипептидаза, гидролизирующий дипептиды, состоящие из остатков L-аминокислот, не действует на такие же дипептиды, состоящие из остатков D-аминокислот. Специфичность действия Ф. послужила нем. учёному Э. Фишеру основанием для сравнения субстрата и Ф., который катализирует его превращение, с замком и соответствующим ему ключом. Стереохимическая специфичность Ф. теснейшим образом связана с одной из основных особенностей живых организмов – их способностью к синтезу оптически активных органических соединений.
В образовании соединения между ферментом и субстратом – т. н. фермент-субстратного комплекса – принимают участие лишь некоторые функциональные группы молекулы Ф., образующие его активный центр . Так, например, в молекуле гидролизирующего белки химотрипсина , состоящего из 246 аминокислотных остатков, активный центр образован одним из остатков серина (химотрипсин относится к сериновым протеиназам) и двумя остатками гистидина, расположенными в удалённых друг от друга участках полипептидной цепи. Сближение этих функциональных групп активного центра происходит благодаря свойственной молекуле химотрипсина специфической пространственной (третичной) структуре. Её нарушение в результате денатурации белка или каких-либо химических модификаций приводит к изменению или полной потере каталитической активности. В случае двухкомпонентных Ф. в образовании фермент-субстратного комплекса принимают участие не только функциональные группы апофермента, но и простетическая группа. Так, при расщеплении пировиноградной кислоты пируватдекарбоксилазой субстрат связывается с частью молекулы тиамин-пирофосфата следующим образом:
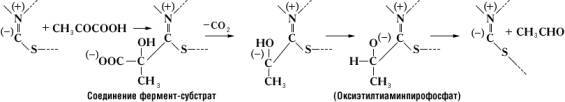
Исключительно высокая специфичность действия Ф. объясняется их белковой природой. Так, пиридоксалевые Ф., содержащие один и тот же кофермент (пиридоксальфосфат), могут принадлежать к различным классам и катализировать самые разнообразные реакции. Специфичность их действия зависит от природы апофермента.
Условия действия ферментов. Действие Ф. зависит от ряда факторов, прежде всего от температуры и реакции среды (pH). Оптимальная температура, при которой активность Ф. наиболее высока, находится обычно в пределах 40–50 °С. При более низких температурах скорость ферментативной реакции, как правило, снижается, а при температурах, близких к 0 °С, практически реакция полностью прекращается. При повышении температуры выше оптимальной скорость ферментативной реакции также снижается и, наконец, полностью прекращается. Снижение интенсивности действия Ф. при повышении температуры сверх оптимальной объясняется главным образом начинающимся разрушением (денатурацией) входящего в состав Ф. белка. Поскольку белки в сухом состоянии денатурируются значительно медленнее, чем белки оводнённые (в виде белкового геля или раствора), инактивирование Ф. в сухом состоянии происходит гораздо медленнее, чем в присутствии влаги. Поэтому сухие споры бактерий или сухие семена могут выдержать нагревание до гораздо более высоких температур, чем те же споры или семена в увлажнённом состоянии.
Важнейшим фактором, от которого зависит действие Ф., как установил впервые С. Сёренсен , является активная реакция среды – pH. Отдельные Ф. различаются по оптимальной для их действия величине pH. Так, например, пепсин, содержащийся в желудочном соке, наиболее активен в сильнокислой среде (pH 1–2); трипсин – протеолитический Ф., выделяемый поджелудочной железой, имеет оптимум действия в слабощелочной среде (pH 8–9); оптимум действия папаина – протеолитического Ф. растительного происхождения – находится в слабокислой среде (pH 5–6).