Для выявления сформированных межсистемных отношений была зарегистрирована активность нейронов лимбической коры в дефинитивном поведении обученных таким образом животных. Из всей совокупности зарегистрированных нейронов для анализа были отобраны клетки, специализированные относительно сформированных обучением в экспериментальной клетке поведенческих актов.
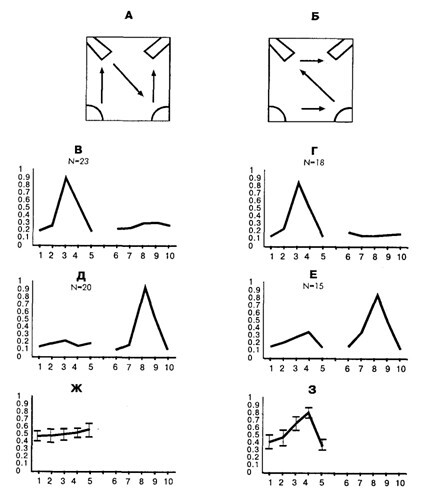
Рис. 15.2 . Усреднённые паттерны активности нейронов, специализированных относительно подхода и нажатия педали в зависимости от времени формирования и стратегии обучения
А, Б – схемы применявшихся стратегий обучения (в нижних углах – кормушки, в верхних – педали); В–З: усреднённые паттерны нейрональной активности. По оси абсцисс – номера поведенческих актов; по оси ординат – нормализованная по максимальной частота активности; В, Г – усреднённые паттерны активности нейронов, специализированных относительно оперирования с первой по порядку обучения педалью; Д, Е– усреднённые паттерны активности нейронов, специализированных относительно оперирования со второй по порядку обучения педалью; Ж, 3 – паттерны (средняя ± стандартная ошибка) неспецифической активности нейронов, представленных на частях Д и Е, но нормализованной по максимуму неспецифической активности. Под схемой конкретной стратегии расположены паттерны активности нейронов, которые зарегистрированы у кроликов, обученных по этой стратегии.
Критерием специализации служило наличие активации нейрона в соответствующем акте во всех случаях его реализации. Как было показано А. Г. Горкиным и Д. Г. Шевченко [1990], этот формальный критерий позволяет достаточно надёжно выделять специфическую связь активности нейрона с конкретным актом поведения. Для анализа неспецифической активности были построены паттерны активности каждого из исследовавшихся нейронов во всех актах пищедобывательного поведения. Паттерн активности представляет собой распределение средних частот импульсации нейрона в выделенных актах поведения. В поведении на одной стороне клетки было выделено 5 актов: поворот головы к педали, перенос лап к педали, нажатие педали, перенос лап к кормушке и захват пищи в кормушке. Всего на двух сторонах клетки было выделено 10 актов. Усреднённые паттерны активности нейронов, специализированных относительно подхода к педали, представлены на рис. 15.2 под схемами соответствующих стратегий обучения. Так как кролик подходил к двум педалям, и паттернов – два для каждой из стратегий. Один из них показывает распределение активности нейронов, специализированных относительно подхода к первой по порядку обучения педали, а другой – относительно второй. Легко заметить, что паттерны у кроликов, обученных по стратегии «Б», различаются для первой и второй педали, в то время как для животных, обученных по стратегии «А», – не различаются. Сходные данные были получены для другой группы специализаций – нейронов, связанных с подходом к кормушке [Горкин, Шевченко, 1995].
Данные, полученные в этой работе, показали, что для систем, не связанных логикой последовательного поведения в одну группировку, их взаимные отношения зависят от истории формирования поведения. Эта зависимость проявляется в виде повышенной степени актуализации второй из двух последовательно формировавшихся систем при реализации первой. В этом исследовании были также выявлены отношения систем, основанные на факторах сходства движений и целей соответствующих поведенческих актов.
В связи с тем, что актуализация функциональных систем, оказалась чувствительной к действию множества факторов, оказывается возможным рассмотреть с позиций системной психофизиологии огромный объем экспериментальных данных, полученных в нейрофизиологии обучения (см. например [Рабинович, 1975; Котляр, 1989]). В основном эти данные касаются пластичности нейрональных ответов на стимулы при выработке условных рефлексов. Сразу стоит обратить внимание на то, что достоверные изменения частоты постстимульной импульсации нейронов в ряду сочетаний, как правило, не превышают 1,5-кратный уровень «фоновой» активности. Это означает, что в случае повышения в обучении возбудительного ответа регистрируется активация, которая в соответствии с упомянутыми ранее критериями не может считаться специфической. Этот факт, вместе с плавной динамикой ответа и наличием тормозных реакций у многих нейронов, свидетельствует, на наш взгляд, о том, что основная феноменология пластичности попадает в разряд изменения межсистемных отношений уже имевшихся до обучения элементов индивидуального опыта. Это вполне соответствует теоретическим положениям бихевиористов о том, что при выработке классических рефлексов и дифференцировок новой реакции, т.е. в терминах данной главы, нового акта поведения не формируется.
* * *
Изложенные ранее теоретические положения и экспериментальные данные позволяют в общих чертах описать основные моменты процесса научения с точки зрения системной психофизиологии.
В дефинитивном поведении циклически повторяющиеся соотношения организма со средой в виде адаптивных последовательностей поведенческих актов формируют отношения элементов опыта. За счёт воздействия среды эти последовательности могут быть прерваны. В соответствии с имеющимся опытом вместо запланированного акта в таком случае реализуется ориентировочное поведение, сменяющееся следующим приспособительным актом. При многократном повторении такого воздействия происходит привыкание, т.е. сокращение перерыва в последовательности вплоть до его полного исчезновения. В случае когда «информационное» воздействие среды сопряжено с «подкрепляющим» в виде разрушающего воздействия либо появляющейся возможности удовлетворения «мотивированного» состояния, после ориентировочного совершается поведенческий акт, не входивший ранее в эту последовательность. При многократном повторении фиксируется новая последовательность смены поведенческих актов и обеспечивающие её осуществление отношения элементов опыта.
Собственно научение, заключающееся в формировании нового элемента индивидуального опыта, начинается с возникновения проблемной ситуации, когда организм не может достигнуть желаемого результата за счёт использования имеющихся в опыте функциональных систем. На нейрональном уровне этому соответствует длительное рассогласование метаболических потребностей клеток и синаптического притока (см. гл. 14). Это приводит к одновременной актуализации множества функциональных систем. Такая актуализация приводит в поисковом поведении к новым последовательностям поведенческих актов и изменениям набора прасистем, т.е. пробным соотношениям организма со средой. За счёт имеющегося опыта поведения индивида в проблемных ситуациях этот набор не случаен, а подчинён опыту генерации успешных проб, накопленному индивидом. В случае неудачи состояние повышенной актуализации множества систем сохраняется и происходит генерация следующей пробы. В случае же успеха полученный результат снижает общий уровень актуализации систем, видимо, за счёт частичного удовлетворения метаболических потребностей нейронов. После ряда успешных проб происходит исключение лишних конкурирующих альтернатив и окончательно складывается новая функциональная система поведенческого акта. Она имеет вид определённой организации актуализированных ранее сформированных систем и «добавки» группы специализированных нейронов, представляющих вновь сформированный элемент опыта в памяти организма. Эта «добавка» обеспечивает консолидацию кооперативного ансамбля, необходимого для достижения данного результата. Одновременно в силу включения нового акта в последовательность имевшихся поведенческих актов формируются отношения нового элемента опыта с уже имевшимися в памяти организма.
Глава 16 СВЯЗАННЫЕ С СОБЫТИЯМИ ПОТЕНЦИАЛЫ МОЗГА (ССП) В ПСИХОФИЗИОЛОГИЧЕСКОМ ИССЛЕДОВАНИИ
1. ОПРЕДЕЛЕНИЕ, ОСНОВНЫЕ ПРОБЛЕМЫ И КРАТКАЯ ИСТОРИЯ МЕТОДА ССП
1.1. Связанные с событиями потенциалы мозга
ССП представляют собой широкий класс электрофизиологических феноменов, которые специальными методами выделяются из «фоновой», или «сырой», электроэнцефалограммы (ЭЭГ). Термин ССП – калька введённого Г. Воном термина Event-Related Potentials of the Brain, ERPs [Vaughan, 1969]. В характеристиках ССП проявляется связь активности мозга с событиями во внешней среде (например, с предъявлением стимулов), во внешне наблюдаемом поведении испытуемого (например, с двигательной активностью) и с психологическими характеристиками активности испытуемого (например, с ожиданием или с принятием решения) [Rockstroh et al., 1982]. «Промежуточное» положение ССП (по выражению Т. Пиктона и Д. Стасса [Picton, Stuss, 1980]), с одной стороны, отражающих активность мозга, а с другой, – характеристики поведения и психологическую феноменологию, обладает очевидной привлекательностью для психофизиологов, поскольку может обеспечить экспериментальные основания для решения фундаментальных проблем психофизиологии (см. для обзора [Psychophysiol. Brain Res., 1990; Rohrbaugh et al., 1990; Event-Related, 1991]).