Принцип динамичности предполагает нестабильность самой системы во времени. Нестабильность определяется текущими изменениями функциональной значимости образующих систему мозговых структур в ходе реализации энграммы. Топография системы, обеспечивающей воспроизведение, меняется от момента к моменту. Мы предполагаем, что эти изменения связаны с достижением максимума активности энграммами иных элементов, расположенных в других структурах. Постоянная смена активностей следа памяти на разных элементах системы является причиной постоянного «блуждания» активных мнестических центов.
Нейрофизиологические исследования распределенности энграммы . В опытах на изолированных нейронах виноградной улитки обнаружены клетки, у которых формирование следа памяти происходит во время ассоциативного обучения, так что после определённого числа сочетаний условного и безусловного стимула формируется энграмма, достигающая уровня актуализации по электрофизиологическим показателям (рис. 6.1). Количество таких нейронов относительно невелико – менее 15% от общего количества зарегистрированных в ситуации ассоциативного обучения (687 нейронов). Более 80% клеток продемонстрировали феномен отсроченного обучения – он заключался в том, что во время предъявления ассоциированных стимулов ответ на «условный» стимул или не изменялся, или же ухудшался при любой частоте предъявления пары. Основная особенность заключалась в том, что увеличение ответа на «условный» стимул после обучения развивалось постепенно (рис. 6.2). [Grechenko, 1993]. Достижение максимальной величины ответа, которая зависит от количества предъявленных сочетаний и от количества проведённых циклов обучения, у разных клеток происходит через неодинаковое время. После выполнения первой серии, состоявшей из предъявления 15–20 пар ассоциированных стимулов, время достижения максимальной величины ответа на условный стимул составляло от 5 до 40 мин (см, рис. 6.2). Опыты на идентифицированных нейронах показали, что независимо от вида ассоциируемых стимулов и от особенностей предъявления сочетаний данный конкретный нейрон всегда обучается по одному и тому же способу – или во время обучения, или отсроченно. Это качество является его индивидуальной характеристикой в отношении данного вида обучения. (В опытах использовали ассоциации стимулов, адресованных различным структурам клетки, – два внутриклеточных деполяризационных стимула, активирующих пейсмекерный механизм, или электровозбудимые мембраны и две микроаппликации медиатора, или микроаппликацию медиатора в комбинации с электрическим стимулом.) По-видимому, в основе этого феномена лежат особенности внутриклеточных процессов, опосредующих ассоциативное обучение, и эти процессы различны по скорости своего развития.
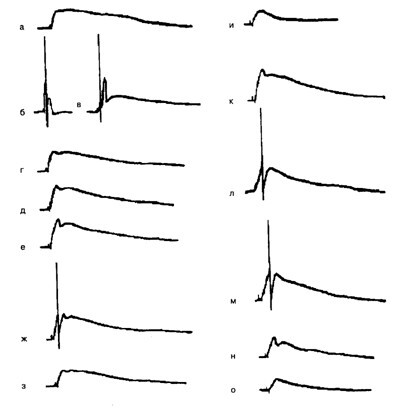
Рис. 6.1 . Формирование условного ответа при сочетании микроаппликации ацетилхолина (АХ) с электрическим деполяризационным импульсом тока (0,36 нА, 100 мс):
а– исходный ответ на микроаппликацию АХ (УС) в локус 1; б – ответ нейрона на деполяризационный стимул, используемый в качестве подкрепления (БС). Сила тока 0,36 нА, длительность 100 мс; в – сочетание АХ и БС, интервал от начало микроаппликации до начала действия БС 50 мс, частота предъявления сочетаний 1 раз в 2–3 мин; Г– ответ нейрона на изолированное предъявление АХ после 8 сочетаний УС–БС; Д – ответ нейрона после 12 сочетаний; е – ответ нейрона на АХ после 20 сочетаний; ж, з – ответ нейрона через соответственно 5 и 25 мин после предъявления 20 сочетаний; н – ответ нейрона на микроаппликацию АХ в локус 2 (ответ получен до проведения обучения в локусе 1); к– ответ нейрона на АХ после 8 сочетаний УС–БС во второй серии; л– ответ нейрона на АХ в локусе 1 после 20 сочетаний во второй серии; м, н – ответ нейрона на АХ через соответственно 30 и 40 мин после обучения; о – ответ нейрона на АХ в локусе 2 после достижения максимального ответа в локусе 1(л). Калибровка: 10 мВ, 1 с
Время сохранения следа памяти в состоянии наивысшей активности на изолированных нейронах не слишком велико – так, после выполнения первой серии обучения у клеток, обучающихся во время предъявления ассоциированных стимулов, оно не превышает 20 мин, а у отсроченно обучающихся нейронов – 40 мин. Время достижения максимума ответа после выполнения второй и последующих серий обучения изменяется. У нейронов первой группы все события развиваются традиционно – при выполнении каждой следующей серии требуется всё меньшее количество ассоциированных стимулов, а время сохранения следа на максимальном уровне актуализации увеличивается (после выполнения 2–4 серий оно может достигать 90 мин). У нейронов же второй группы выполнение каждой следующей серии значительно продлевает время «жизни» следа – после второй серии оно может увеличиться в 2 раза – и, как ни удивительно, увеличивает время достижения максимальной активности следа памяти. Например, если след после первой серии обучения достигал наиболее высокого уровня актуализации через 10 мин, то после второй или третьей серии – только через 30–40 мин. Кажется вероятным, что такие характеристики пластичности нейронов могут лежать в основе распределенности энграммы по популяции клеток, опосредующих конкретную форму поведения. Воспроизведение следа памяти через разное время после обучения происходит с различных нейронов, отличающихся временными характеристиками достижения максимальной активности, инициированной обучением.
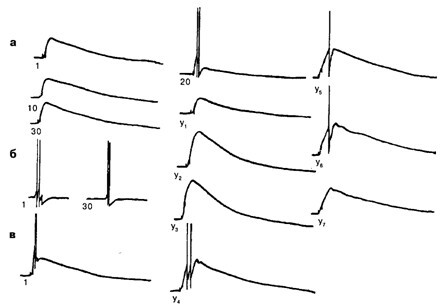
Рис. 6.2. Отсроченное обучение на нейроне ЛПаЗ при сочетании микроаппликаций АХ с электрическим деполяризационным стимулом, вызывающим генерацию ПД:
а – исходный ответ нейрона на микроаппликацию АХ (1), ответ на АХ при повторных микроаппликациях на 10-е и 30-е применение. Частота микроаппликаций 1 раз в 2–3 мин; б – ответ нейрона при повторных предъявлениях БС с частотой 1 раз в 2–3 мин. Сила стимула 2,5 нА, длительность 100 мс: показан ответ нейрона на 1-е и на 30-е предъявление; в – формирование условного ответа при сочетании АХ с электрическим предъявлением. Интервал между УС и БС равен 10 мс: 1 – ответ нейрона на первое сочетание УС–БС; 20 – на 20-е предъявление УС–БС; Y1 – ответ нейрона на АХ через 3 мин после 20 сочетаний; У2 У3 У4, У5 У6 Уz - ответ нейрона через 10, 30, 40, 90, 120 и 150 мин после пятой серии сочетаний соответственно. Калибровка: 10 мВ, 1 с
Факты, полученные в опытах на изолированных нейронах, совпадают по существу с данными экспериментов, проведённых на полуинтактном препарате улитки [Максимова, Балабан, 1983]. В частности, для командного идентифицированного нейрона ЛПаЗ в этих опытах получили весьма похожие временные параметры актуализации активной энграммы – около 90 мин после выполнения трёх серий предъявлений ассоциированных стимулов. (В опытах изучали условную пассивно-оборонительную реакцию.) Так как на поведенческом уровне этот условный ответ обнаруживается непосредственно после обучения, то, следовательно, он осуществляется ансамблем нервных клеток, в котором не участвует командный нейрон ЛПаЗ (его энграмма актуализируется отсроченно). Это наблюдение заставляет предполагать, что в зависимости от времени, прошедшего после обучения, реализацию следа памяти осуществляют разные по своему составу нейронные ансамбли. Изменение элементов системы обеспечивает функциональную неоднородность энграммы, воспроизводимой через разное время после обучения.
4. ПРОЦЕДУРНАЯ И ДЕКЛАРАТИВНАЯ ПАМЯТЬ
В последнее время стало приобретать всё большее значение представление о множественности систем памяти. Это представление сформировалось на основе данных, полученных при исследовании больных с различными поражениями мозга, а также в опытах на здоровых испытуемых, выполненных с использованием регистрации вызванных потенциалов, и в опытах на животных с различными повреждениями мозговых структур.
Эти системы памяти имеют разные оперативные характеристики, участвуют в приобретении знаний разного рода и осуществляются разными мозговыми структурами. Исследователи предположили, что переработка по крайней мере двух видов информации ведётся в мозгу раздельно и каждый из этих видов хранится также отдельно [Squire, 1994]. Упомянутые ранее данные, полученные как на амнезированных пациентах, так и на людях с обычной памятью и на животных, позволили разделить системы памяти на две большие группы; процедурную и декларативную память.