Ранее уже говорилось о том, что влияние эмоций на поведение определяется отношением животного к своему эмоциональному состоянию, причём подчинено принципу максимизации положительных эмоций и минимизации отрицательных. Этот принцип реализуется влиянием мотивационно-эмоциональных структур гипоталамуса на информационные (когнитивные) и организующие движение отделы новой коры, о чём свидетельствует анализ пространственной синхронизации электрической активности мозговых структур при самостимуляции крыс слабым постоянным током [Pavlygina et al., 1976].
На рис 7.2 отображён процент случаев регистрации значимой когерентности (Р < 0,05) на частотах альфаи тета-диапазонов в потенциалах сравниваемых мозговых образований. Рассматривая рисунок, можно убедиться, что непосредственно перед нажатием на педаль резко (более чем в 3 раза) увеличивается процент случаев статистически значимой когерентности электрической активности эмоционально позитивного пункта гипоталамуса с моторной областью коры. Возросшая когерентность свидетельствует о готовности нервных путей к проведению возбуждения по трём каналам условных связей: от ранее раздражавшегося эмоционально позитивного пункта к двигательной коре, к зрительной коре, а также между зрительным анализатором, воспринимающим условный сигнал будущего подкрепления (вид педали, её местонахождение в камере и т.п.), и двигательной областью коры, поскольку именно вид педали будет направлять движение животного, инициированное следовым возбуждением эмоционально позитивной зоны.
В период пребывания крысы на педали когерентность уменьшается, животное получает подкрепление и становится совершенно пассивным. Непосредственно перед уходом впервые увеличивается когерентность негативного пункта и моторной области коры: возбуждение негативных структур готово к трансформации в двигательную реакцию избавления. После ухода животного с педали процент случаев достоверного увеличения когерентности возвращается к исходным цифрам. Только следы эмоционально негативного состояния всё ещё обнаруживаются при сопоставлении электрической активности негативного пункта с двигательной областью коры.
Как показали исследования Р.А. Павлыгиной и Ю.В. Любимовой [1994], мотивационные влияния гипоталамуса на новую кору носят асимметричный характер. Эта асимметрия проявляется после односуточной пищевой депривации кроликов не только в преимущественной активации левого полушария, но и в усилении взаимосвязи электрических процессов левого полушария. Анализ спектрально-корреляционных характеристик электрической активности мозга кролика обнаружил большую реактивность левого полушария по сравнению с правым и при состоянии жажды [Маликова, Мац, 1991]. Преобладающая активность левого полушария при возникновении голода у человека зарегистрирована в альфаи дельта-диапазонах его электроэнцефалограммы [Сулимов, 1995].
О межполушарной асимметрии при естественной пищевой мотивации свидетельствует регистрация импульсной активности отдельных нейронов зрительной и сенсомоторной области неокортекса кроликов, а также взаимодействие этих нейронов. Судя по импульсации нейронов, кора левого полушария более активна у голодных кроликов, а кора правого полушария – у сытых. Наиболее выраженные различия наблюдались в активности нейронов фронтальных областей, наименее выраженные – в сенсомоторных нейронах [Павлова, 1995]. В ходе этих исследований была впервые установлена функциональная асимметрия самого «пищевого» гипоталамуса. При стимуляции левого гипоталамуса пороги пищевых реакций были меньше, а сами реакции более стабильны, чем при стимуляции правого гипоталамуса, причём «пищевая» асимметрия латерального гипоталамуса не коррелировала с моторной и сенсорной асимметрией у этих животных [Павлова, Мац, 1996].
Вся совокупность имеющихся в настоящее время данных свидетельствует о том, что гипоталамус является ключевой структурой для реализации наиболее древней подкрепляющей функции эмоций, для решения универсальной поведенческой задачи максимизации-минимизации возникшего эмоционального состояния: приближения или избегания. Ведь именно получение желательных, эмоционально положительных стимулов или устранение нежелательных, эмоционально отрицательных, а не удовлетворение какой-либо потребности служит непосредственным подкреплением при обучении. У крыс невозможно выработать инструментальный условный рефлекс при введении пищи через канюлю в желудок (т.е. минуя вкусовые рецепторы), хотя такой рефлекс вырабатывается при введении в желудок морфина, который очень быстро вызывает у животного положительное эмоциональное состояние. Тот же морфин из-за своего горького вкуса перестаёт быть подкреплением, если его вводить через рот [Cytawa, Trojniar, 1976]. В другой серии опытов авторы вырабатывали инструментальный пищевой условный рефлекс у крыс, а после его упрочения заменяли натуральную пищу введением питательного раствора в желудок через носоглоточную канюлю. Рефлекс нажатия на рычаг при этом угасал, но сохранялся, если в желудок вводили 0,05%-ный раствор морфина [Trojniar, Cytawa, 1976].
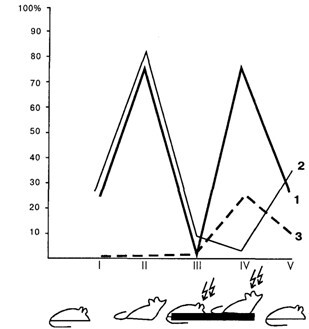
Рис. 7.2. Процент случаев статистически значимого изменения когерентности электрической активности мозговых структур:
/– спокойное состояние; //– перед посадкой на педаль; ///– на педали; IV– перед уходом с педали; V – сразу же после ухода; 1 – эмоционально положительный пункт гипоталамуса – двигательный неокортекс; 2– эмоционально отрицательный пункт гипоталамуса – двигательный неокортекс; 3 – двигательная область – зрительная область неокортекса
В отличие от гипоталамуса, сохранность второго «мотивационного» мозгового образования – миндалины – необязательна для формирования сравнительно простых форм временных связей. По данным М.Л. Пигаревой [1983], двухсторонняя амигдалоэктомия не препятствует выработке пищевых и оборонительных условных рефлексов. При этом миндалина «безразлична» к вероятности подкрепления, к его информационному компоненту. Скорость выработки условных двигательных пищевых рефлексов у амигдалэктомированных крыс не отличается от аналогичных показателей у интактных животных ни при высокой (100 и 50%), ни при низкой (25%) вероятности их подкрепления. Только вероятность в 33%, для которой характерен максимум возникающего при этом эмоционального напряжения, сопровождается некоторым замедлением процесса формирования пищедобывательной реакции. Разрушение миндалин у крыс через неделю после однократного обучения реакции избегания не препятствует воспроизведению этого навыка через четыре дня после операции [Parent et al., 1995]. При одностороннем выключении правой или левой миндалины через 24 ч или 10 дней после сеанса выработки реакции избегания оказывается, что время задержки реакции короче в том случае, если была выключена правая миндалина [Coleman-Mesches, McGaugh, 1995].
Результаты опытов на животных совпадают с клиническими наблюдениями. У двух амигдалэктомированных пациентов не отмечено нарушений формирования ассоциаций между объектами двух разных сенсорных модальностей [Lee et al., 1995]. Двухстороннее поражение миндалин не приводит к амнезии. Дефект памяти носит строго избирательный характер, связанный с эмоциональной окраской событий [Markowitsch et al., 1994].
Сохранность миндалины наиболее существенна в том случае, когда возникает конкуренция между сосуществующими мотивациями и становится необходимым выбор доминирующей потребности, подлежащей удовлетворению в данный момент. Хорошей экспериментальной моделью подобной ситуации может служить выработка условно-рефлекторного переключения разнородных условных рефлексов по Э.А. Асратяну [Пигарева, 1983], когда один и тот же сигнал (звук) утром подкрепляется пищей, а вечером – болевым раздражением. Судя по проценту правильных оборонительных и пищевых условных реакций, на протяжении 40 дней не удалось выработать условнорефлекторное переключение у амигдалэктомированных крыс. Вместе с тем решение подобной поведенческой задачи возможно, если искусственно создать достаточный дисбаланс между конкурирующими мотивациями и соответствующими им эмоциями: между голодом и страхом (табл.).