Таблица 5.5. Расстояния между группами; FM-алгоритм, шаг 1a
.31 .93
.863
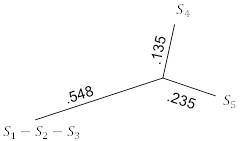
Имея только три таксона в этой таблице, можем точно подогнать данные к дереву, используя 3-точечные формулы, чтобы получить рисунок 5.10. Ключевым моментом здесь является то, что 3-точечные формулы, в отличие от UPGMA, могут давать неравные расстояния таксонов от общего предка.
Рисунок 5.10. FM-алгоритм; шаг 1.
Теперь оставляем только ребра, заканчивающиеся в
и
на рисунке 5.10, и возвращаемся к исходным данным. Помните, что группа
была нужна только временно, чтобы могли использовать 3-точечные формулы; пока не собирались объединять эти таксоны. Однако, поскольку объединили
и
, объединяем их в группу для остальной части алгоритма, как сделали бы с UPGMA. Это формирует таблицу 5.6.
Таблица 5.6. Расстояния между группами; FM-алгоритм, шаг 1b
1.005 .72 .965
.61 .42
.37
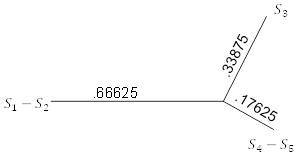
Снова ищем ближайшую пару (теперь это
и
) и соединяем их аналогичным образом. Объединяем все, кроме
и
, в одну временную группу
и вычисляем расстояния
и
. Полученными значениями заполняем таблицу 5.7. Применение трехточечной формулы к таблице 5.7 дает рисунок 5.11.
Таблица 5.7. Расстояния между группами; FM-алгоритм, шаг 2a
.683 .783
.37
Рисунок 5.11. FM-алгоритм; шаг 2.
Оставляем ребра инцидентные с
и
на рисунке 5.11, отбрасывая ребро, ведущие к временной группе
. Таким образом, теперь есть две объединенные группы,
и
. Чтобы вычислить новую таблицу, содержащую эти две найденные группы, усредняем расстояния
и
. Выше уже вычислили
, поэтому получаем таблицу 5.8.
Таблица 5.8. Расстояния между группами; FM-алгоритм, шаг 2b
1.005 .8425
.515
На этом этапе можем получить итоговое дерево по таблице путем окончательного применения 3-точечных формул, что дает рисунок 5.12.
Рисунок 5.12. FM-алгоритм; шаг 3.
Теперь заменяем группы на этой последней диаграмме шаблонами ветвления, которые уже нашли ранее. Это дает рисунок 5.13.
Последним шагом является заполнение оставшихся длин
и
, используя длины, показанные на рисунке 5.12. Так как
и
в среднем дают расстояние
от соединяющей их вершины, а
и
находятся в среднем на
от соединяющей их вершины, то
и
получаем для присвоения длин оставшимся ребрам.