Таким образом, молекулярные часы означают, что количество мутаций на любом ребре пропорционально прошедшему времени, при этом константа пропорциональности представляет собой постоянную скорость мутации. Если предположить, что существуют молекулярные часы, то независимо от того, рисуем ли длины ребер, представляющие количество мутаций или только прошедшее время, то нарисуем одну и ту же фигуру с точностью до масштаба этой константы.
Если гипотеза молекулярных часов справедлива для корневого метрического дерева, то каждый лист будет расположен на одинаковом общем расстоянии от корня дерева. Это связано с тем, что расстояния от корня пропорциональны времени, прошедшему с тех пор, как таксоны начали расходиться с общим предком. У каждого таксона было одинаковое количество времени, чтобы эволюционировать от корневого предка, поэтому каждый таксон накопит одинаковое количество мутаций.
Без молекулярных часов связь между количеством мутаций вдоль ребра и количеством времени может быть сложной для моделирования. Предположим, что вдоль одного ребра филогенетического дерева частота мутаций была довольно мала, а вдоль другого – частота мутаций была большой. Затем несмотря на то, что оба края могут соответствовать одинаковому количеству времени, вдоль одного из них произойдет значительно больше мутаций. Без получения какой-либо дополнительной информации о скорости мутации – возможно, путем сравнения с летописью окаменелостей – обычно нет способов определения прошедшего времени, связанного с ребрами деревьев.
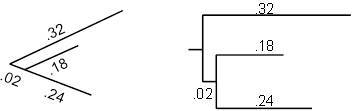
Метрические деревья иногда рисуются в «квадратном» стиле, чтобы было легче сравнивать расстояния по различным эволюционным путям. Например, два дерева на рисунке 5.4 представляют одну и ту же информацию. В дереве слева ребра имеют указанную длину, а в дереве справа горизонтальные ребра имеют те же длины. Таким образом, вертикальные ребра на правом дереве считываются как не вносящие никакого вклада в количество мутаций; они служат исключительно для разделения различных линий для повышения читабельности.
Рисунок 5.4. Разные изображения одного и того же метрического дерева.
Задачи для самостоятельного решения:
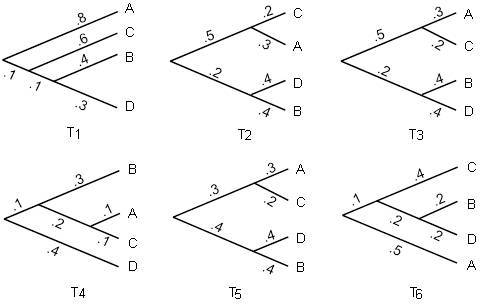
5.1.1. Рассмотрим деревья на рисунке 5.5.
Рисунок 5.5. Деревья для задачи 5.1.1.
а. Какие из них совпадают с корневыми метрическими деревьями?
б. Какие из них совпадают с некорневыми метрическими деревьями?
в. Какие из них такие же, как корневые топологические деревья?
г. Какие из них такие же, как некорневые топологические деревья?
д. Для каких деревьев работают молекулярные часы?
5.1.2. а. Нарисуйте единственное топологически уникальное некорневое раздвоенное дерево, которое могло бы описать связь между 3 таксонами.
б. Нарисуйте три топологически различных корневых раздвоенных дерева, которые могли бы описать связь между 3 таксонами.
5.1.3. а. Нарисуйте все 3 топологически различных некорневых раздвоенных деревьев, которые могли бы описать связь между 4 таксонами.
б. Нарисуйте все 15 топологически различных корневых раздвоенных деревьев, которые могли бы описать связь между 4 таксонами.
5.1.4. Для
терминальных таксонов количество некорневых раздвоенных деревьев можно найти как
. Составьте таблицу значений и отобразите эту функцию для
.
5.1.5. Для
терминальных таксонов количество корневых раздвоенных деревьев равно значению
. Составьте таблицу значений и отобразите эту функцию для
.
5.1.6. В этой задаче рассмотрим рассуждения, лежащие в основе формул для числа топологически различных деревьев, корневых и некорневых.
а. Предположим известно, что некорневое дерево с
концевыми вершинами состоит из
ребер. Объясните, почему некорневое дерево с
концевыми вершинами будет иметь
ребра. Подсказка: подумайте о том, как добавление еще одной конечной вершины в существующее дерево влияет на количество ребер.
б. Поскольку некорневое дерево с 2 концевыми вершинами имеет 1 ребро, объясните из пункта (а), почему некорневое дерево с
концевыми вершинами будет иметь
ребра.
в. Предположим известно, что существует
некорневых деревьев с
концевыми вершинами. Объясните, почему существует
некорневых деревьев с
концевой вершиной. Подсказка: подумайте, сколькими различными способами можно добавить еще одну конечную вершину к существующему дереву.
г. Поскольку существует только 1 некорневое дерево с 2 концевыми вершинами, объясните используя пункт (c), почему существует
некорневых деревьев с
концевыми вершинами при
.
д. Объясните, почему
.
е. Почему число корневых деревьев с
концевыми вершинами такое же, как число некорневых деревьев с
концевыми вершинами?
ж. Сделайте вывод о правильности формул в задачах 5.1.4 и 5.1.5.
5.1.7. Поскольку митохондриальная ДНК у человека наследуется исключительно от матери, она может быть использована для структуры, относящейся к любому количеству людей из разных этнических групп, предполагая, что все люди произошли от одной первой человеческой самки. В зависимости от модели кластеризации этнических групп, это может дать представление о физическом местоположении той женщины, которую иногда называют митохондриальной Евой.
В работе Канна 1987 года была впервые предпринята попытка определить местонахождение митохондриальной Евы в Африке. Поддерживая теорию происхождения человека «из Африки», было построено дерево с корнями, которое, как утверждается, показывает отношения между 147 людьми. Сколько топологически различных деревьев нужно было бы рассмотреть, если бы действительно рассматривалась каждая возможность? Возможно, для ответа на этот вопрос придется использовать формулу Стирлинга:
. Здесь символ «∼» можно интерпретировать как «приблизительно». Изучению последствий трудности рассмотрения стольких деревьев посвятил свою работу Гиббонс в 1992 году.
5.1.8. Филогенез четырех терминальных таксонов A, B, C и D связан по определенному метрическому дереву. Суммарные расстояния между таксонами вдоль дерева оказались такими же, как в таблице 5.1.
Таблица 5.1. Расстояния между таксонами для задачи 5.1.8