Денис Соломатин
Математические модели в естественнонаучном образовании. Том II
Глава 5. Построение филогенетических деревьев
Смоделировав эволюцию ДНК в предыдущей главе, теперь готовы использовать эти модели для важных выводов из реальных данных ДНК. Увидим, как модель молекулярной эволюции вместе с некоторыми новыми математическими методами может быть использована для восстановления хронологии событий эволюционной истории. Давайте рассмотрим хорошо изученный, но все же удивительный вопрос: кем приходятся люди современным обезьянам? Более точно, какие из горилл, шимпанзе, орангутанов и гиббонов являются нашими ближайшими эволюционными родственниками, или все эти обезьяны более тесно связаны друг с другом, чем с нами?
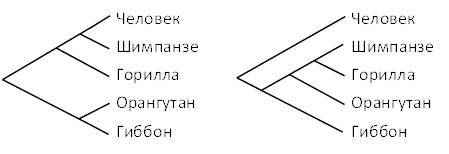
Рисунок 5.1. Две возможные филогении гоминоидов.
Ранние эволюционисты рассматривали шимпанзе и горилл как наших ближайших родственников. Считалось, что люди и эти африканские обезьяны образуют одну эволюционную группу, которая отделилась от других линий обезьян в более отдаленном прошлом. Чуть позже господствующим стало мнение, что все современные человекообразные обезьяны более тесно связаны друг с другом, чем с людьми. Две возможные схемы, представляющие более подробные версии этих конкурирующих взглядов на эволюцию гоминоидов, показаны на рисунке 5.1.
Вопросы для самопроверки:
– Поскольку шимпанзе и горилла являются африканцами, в то время как орангутанг и гиббон являются азиатами, что, если вообще что-либо, каждое из этих деревьев укажет о вероятном месте появления первых людей?
Как выбрать, какое из этих или многих других возможных эволюционных деревьев является лучшим описанием происхождения гоминоидов? Один из подходов заключается в том, что сначала выбирают конкретный ген, общий для всех человекообразных обезьян и людей, но последовательность ДНК которого показывает вариации от вида к виду. Если предположить, что этот ген является общим и сходным для всех гоминоидов, поскольку он произошел от общего предка (то есть последовательности ортологичны, другими словами, к их разделению привел процесс видообразования), то вариации последовательностей среди видов должны содержать информацию об их эволюционной истории.
Например, 898-элементные пары последовательностей митохондриальной ДНК HindIII из этих гоминоидов и семи других приматов были зарегистрированы в работе Хаясака с соавторами 1988 года, он в свою очередь опирался на работу коллективов под руководством Андерсона 1981 года и Брауна 1982 года. Эти последовательности согласуются на 67-97% участков, в зависимости от того, какие из них сравниваются. Чтобы увидеть эти последовательности, загрузите базу данных primatedata в MATLAB, а затем посмотрите имена переменных, в которых они хранятся.
Хотелось бы вывести филогенетическое дерево, подобное одному из представленных на рисунке 5.1, показывающее, как все обезьяны эволюционировали от общего предка. Но какие данные указывают на «лучшее» дерево или даже «хорошее» дерево, чтобы объективно описать эволюционное происхождение?
Конечно, ученые уже рисовали деревья, показывающие подозрительные эволюционные отношения между видами задолго до появления методов секвенирования ДНК. Морфологическое сходство между видами является одним из источников подсказок относительно того, какие деревья адекватно описывают происхождение видов. Идентификация общих предков по окаменелостям – это другой подход. Теперь данные последовательности предоставляют новый источник информации об эволюционной истории, но использование их для вывода филогенетических деревьев требует разработки новых математических инструментов.
5.1. Филогенетические деревья
Прежде чем начнем разрабатывать методы построения филогенетических деревьев, понадобится некоторая терминология. Поскольку последовательности, которые, возможно, захотим связать, могут происходить от разных видов, как в примере с гоминоидами, или вместо этого от разных подвидов, популяций или даже отдельных особей, будем называть каждый источник последовательности ДНК таксоном (множественное число таксоны). Эквивалентный общеупотребительный термин – операционная таксономическая единица, обычно обозначаемая аббревиатурой ОТЕ (в иностранной литературе можно встретить обозначение OTU).
Будет стараться нарисовать диаграмму, состоящую из отрезков линий, которая представляет собой эволюционную историю таксонов. Каждый из сегментов линии на диаграмме по устоявшейся в теории графов терминологии называется ребром. Диаграмма, подобная приведенной выше, в которой нет циклов и петель, образованных ребрами, называется деревом.
Вопросы для самопроверки:
– Почему разумно предположить, что эволюционные отношения могут быть смоделированы путем рисования именно деревьев? Что бы это значило, если бы существовали цикл или петля?
Поскольку существует боковой перенос генов, например, когда вирусная ДНК постоянно включается в ДНК хозяина, деревья не могут описать все эволюционные отношения. Они обеспечивают простейшую модель, которая, тем не менее, полностью адекватна для большинства применений.
Точка, в которой сходятся нескольких ребер называется внутренней вершиной, в то время как висячий конец ребра у таксона называется конечной вершиной или листом дерева. Вершина, в которой будет находиться общий предок всех таксонов, называется корнем.
Говорят, что дерево раздваивается, находится в состоянии бифуркации, если на каждой его внутренней вершине встречаются по три ребра, а у корня сходится два ребра, как на деревьях на рисунке 5.1. Такие деревья называют двоичными или бинарными. Хотя с биологической точки зрения возможно, что дерево, отличное от двоичного, могло бы описывать эволюционную историю, обычно эту возможность игнорируют.
Вопросы для самопроверки:
– Каково было бы эволюционное значение вершины в дереве, где встречаются четыре ребра (то есть, где маршрут из одного ребра расходится на три направления)? Можете ли представить себе правдоподобные обстоятельства, при которых несколько видов могут расходиться таким образом?
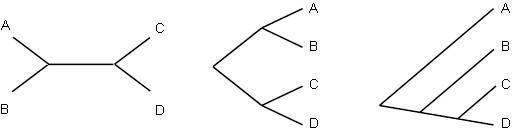
Хотя в идеале каждое филогенетическое дерево должно иметь корень, показывающий общего предка таксонов, иногда приходится обходиться без него. Некоторые методы филогенетического построения деревьев дают некорневые деревья. Например, на рисунке 5.2 показано некорневое дерево и несколько корневых деревьев, которые с ним согласуются. Два дерева справа могли быть согнуты и растянуты, чтобы выглядеть как дерево слева; их отличает только расположение корня.
Рисунок 5.2. Некорневое дерево (слева) и две его корневые версии (в центра и справа).
Посмотрим на деревья с топологической точки зрения. Дерево, относящееся к ряду таксонов, может фактически указывать несколько различных типов информации об их отношениях. Во-первых, если не указываем длины ребер, а значит, смотрим только на ветвящуюся структуру, то рассматриваем только топологию дерева. Считается, что два дерева топологически одинаковы, если можно согнуть и растянуть ребра одного из них, чтобы получить второе дерево. Однако нельзя отрезать ребро и снова прикреплять его в другом месте; это может дать дерево, которое топологически отличается от исходного.
На рисунке 5.3 деревья
,
и
топологически совпадают с некорневыми деревьями, потому что, если бы какая-либо из этих фигур была сделана из резины, ее можно было бы деформировать в другие, не разрезая и не склеивая куски вместе. Дерево
, напротив, топологически отличается от
,
и
.